Gehirntheorie der Wirbeltiere
ISBN
978-3-00-064888-5
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
3.2 Das Nervensystem der segmentierten Bilateria
Aus den einfachen unsegmentierten Bilateria der Replikationsstufe 2 gingen die segmentierten Bilateria der Replikationsstufe 3 hervor. Dies erfolgte über den Zwischenschritt der Koloniebildung durch Knospung mit unterbliebener Abtrennung der Knospe. Jede Knospe bildete ein Körpersegment, anfänglich waren diese Segmente untereinander völlig identisch. Die Ressourcen wurden gemeinsam genutzt. Es gab letztlich einen gemeinsam genutzten Verdauungsapparat, ebenso einen gemeinsamen Kreislauf und später eine gewisse Arbeitsteilung.
Das Nervensystem bestand in jedem Segment zunächst so weiter, wie es beim unsegmentierten Bilateria vorhanden war. Es gab bilaterale sensorische Zentren, motorische Zentren und Mittelwertzentren. Neuronen der Klasse 3 übertrugen die Rezeptorsignale vom sensorischen ins ipsilaterale motorische Zentrum des gleichen Segments. Dortige Motoneuronen steuerten die Zielmuskeln im gleichen Segment an. Es gab in jedem neuronalen Zentrum die laterale Hemmung zur Kontrastverstärkung. Ebenso gab es in jedem Segment die kontralaterale Hemmung, die sowohl auf die Sensorik, die Motorik und die Mittelwertzentren einwirkte. Sowohl das sensorische als auch das motorische Zentrum projizierten ipsilateral in die verschiedenen Mittelwertkerne, von denen eine Rückprojektion zur Vorerregung erfolgte.
Wir beobachten bei real existierenden segmentierten Bilateria einen Signalaustausch zwischen den Segmenten. Ebenso wie das Kreislaufsystem oder das Verdauungssystem segmentübergreifend funktionierte, bildete sich auch neuronal ein segmentübergreifendes Zusammenspiel aus. Es entwickelten sich Projektionsneuronen, die gleichartige neuronale Zentren miteinander verknüpften. Hierbei gab es zwei Signalrichtungen: Eine afferente Projektion war kopfwärts gerichtet, eine efferente dagegen schwanzwärts. Verknüpft wurden auf diese Weise identische Strukturen (Zentren) von zwei benachbarten Segmenten. Wir ordnen den segmentüberschreitenden Projektionsneuronen zwei neue Neuronenklassen zu. Projektionsneuronen, die segmentüberschreitend afferent projizieren, ordnen wir die Neuronenklasse 4 zu. Projektionsneuronen, die segmentüberschreitend efferent projizieren, ordnen wir die Neuronenklasse 5 zu. Hierbei postulieren wir, dass die Neuronen der Klasse 4 die sensorischen Signale übertragen, die Neuronen der Klasse 5 dagegen die motorischen.
Damit gab es bei diesen Tieren insgesamt 6 Neuronenklassen, die als Projektionsneuronen wirkten. Diese sechs Neuronenklassen finden wir noch heute im Gehirn des Menschen wieder, aber auch bei allen Wirbeltieren.
Bei segmentierten Bilateria unterscheiden wir in jedem Segment die Eigensignale von den Fremdsignalen. Eigensignale stammen direkt oder indirekt (über Neuronenketten) von den Rezeptoren dieses Segments ab, egal, ob sie sensorische Signale, motorische Signale oder Signale aus den Mittelwertzentren dieses Segments darstellen.
Fremdsignale entstammen den anderen Segmenten.
Die aufsteigende (afferente) Projektion erfolgt derart, dass der im sensorischen Zentrum ankommende Input - sowohl der von den sensorischen Rezeptoren als auch der von den Mittelwertzentren dieses Segments - dem sensorischen Zentrum des kopfseitigen Nachbarsegments der gleichen Körperhälfte hinzugefügt wird. Hierfür sind Projektionsneuronen der Klasse 4 zuständig. So nimmt die Inputmenge in Kopfrichtung von Segment zu Segment zu, zumal der segmentübergreifende Output an separate, eigenständige Neuronen übergeben wird, so dass die Neuronenanzahl in Kopfrichtung von Segment zu Segment ansteigt.
Hierbei unterstellen wir eine topologische Ordnung hinsichtlich der signalempfangenden Neuronen. Jedes Segment fügt seine Information im übergeordneten Segment so ein, dass sie von Neuronen empfangen wird, die sich seitlich an die bisherigen Neuronen anlagern.
Somit empfängt jedes Segment einen Eigeninput und einen Fremdinput. Der Eigeninput entstammt dem eigenen Segment, also dessen Rezeptoren und neuronalen Kernen. Der Fremdinput entstammt den anderen Segmenten und wurde zu diesem Segment hindurchgereicht.
Das hat den Vorteil, dass im Kopfsegment die komplette sensorische Information über das Gesamtlebewesen vorliegt. Daher bildete sich das Gehirn am kopfseitigen Teil des Tieres.
Was geschah mit dem Input, der in jedem Segment und auf jeder Körperseite in jedem sensorischen Zentrum eintraf?
Zum Ersten wurde er komplett an das benachbarte kopfseitige Segment weitergereicht über Projektionsneuronen der Klasse 4. Nur im Kopfsegment war dies nicht mehr möglich und unterblieb, weil es kein weiteres übergeordnetes Segment gab.
Zweitens wurde der Input komplett an das motorische Zentrum der gleichen Körperseite weitergereicht. Dort angekommen, nahm der Eigeninput einen anderen Weg als der Fremdinput.
Denn der Eigeninput entstammte dem gleichen Segment. Er diente nach wie vor der Ansteuerung der Motoneuronen dieses Segmentes und bewirkte die motorische Antwort in diesem Segment.
Der Fremdinput entstammte den tiefer in Schwanzrichtung zugeordneten Segmenten. Er war vom Herkunftssegment auf Neuronen der Klasse 4 aufgestiegen, hatte zur motorischen Seite gewechselt und nun über Neuronen der Klasse 5 wieder das Herkunftssegment erreicht. Hier dockte er an den zugeordneten Motoneuronen an und bewirkte entsprechende motorische Reaktionen.
So erreichte der sensorische Output eines beliebigen Segments das motorische Zentrum mehrfach. Einmal direkt über Neuronen der Klasse 3, die waagerecht zu ihm zogen. Zum Zweiten wurde er auf den Axonen der Neuronenklasse 4 kopfwärts gesendet, von Segment zu Segment übergeben, in jedem dieser Segmente über die waagerechte Projektion der Klasse 3 zum sensorischen Zentrum geschickt und von dort wieder über Neuronen der Klasse 5 abwärts zum Zielsegment, wo er an den Motoneuronen endete. Je mehr Segmente sich kopfwärts über einem Segment befanden, um so mehr Signalwege gab es für ein sensorisches Signal, um das Zielneuron des zugehörigen Segments zu erreichen.
Somit gab es auf jeder Körperseite einen Nervenstrang, der aus Neuronen der Klasse 4 bestand, die eine afferente Projektion von Segment zu Segment realisierten. Auf der anderen Seite gab es einen Nervenstrang aus Neuronen der Klasse 5, die die efferente Projektion von Segment zu Segment verwirklichten, die zugehörigen Signale waren motorischer Art.
Stellt man sich das Tier so vor, dass sich oben der Kopf und unten der Schwanz befindet, so gab es in jedem Segment eine waagerechte Projektion vom sensorischen zum motorischen Zentrum, die von den Neuronen der Klasse 3 bewirkt wurde. Daher hatte dieses Nervensystem einer Körperhälfte das Aussehen einer Strickleiter.
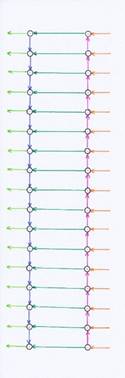
Abbildung 1- einfaches Strickleiternervensystem ohne Mittelwertzentren
Erläuterungen zur Abbildung 1:
Von links trifft in jedem Segment der Input der Rezeptoren ein und erreicht das sensorische Zentrum jedes Segments.
Er wird von den Axonen der Neuronen der Klasse 3 waagerecht zum motorischen Zentrum übertragen.
Weiterhin übertragen Axone der Neuronen der Klasse 4 diese Erregung auch in das übergeordnete Segment.
Die Axone der Klasse 5 im motorischen Zentrum jedes Segments übertragen die eintreffende Erregung einerseits zu den Motoneuronen des Segments, andererseits projizieren sie ins motorische Zentrum des untergeordneten Segments.
Die Mittelwertzentren sind in dieser Darstellung wegen der Übersichtlichkeit weggelassen.
Jede Körperhälfte verfügt über ein eigenes Strickleiternervensystem. Beide sind über Kommissuren miteinander verbunden.
Dieses Strickleitersystem gab es wegen der Bilateralität doppelt, jeweils einmal auf der linken und einmal auf der rechten Körperhälfte. Diese zwei Strickleitersysteme waren jedoch nicht unabhängig voneinander. In jedem Segment gab es die Projektion der Neuronen der Klasse 2, die einerseits das motorische Zentrum einer Seite mit der anderen Seite verknüpften, dort an hemmenden Interneuronen endeten und eine laterale Hemmung zur Kontrastverstärkung realisierte. So standen alle motorischen Zentren einer Körperhälfte mit der anderen in Konkurrenz. Aber auch die sensorischen Zentren waren so verknüpft. So steht jede neuronale Struktur einer Körperseite mit der gleichen neuronalen Struktur der Gegenseite (im gleichen Segment) in neuronaler Konkurrenz. Dies betrifft auch die Signale aus den Mittelwertzentren.
Daher besaß das Gesamtlebewesen ein aus vier Nervensträngen bestehendes Nervensystem, welches in jedem Segment durch querverlaufende Kommissuren der Neuronenklasse 3 und durch querverlaufende Kreuzkommissuren der Neuronenklasse 2 verknüpft waren. Dieses wird als tetraneurales Nervensystem bezeichnet.
Bezüglich der Mittelwertzentren lässt sich der Signalverlauf einfach beschreiben. Jedes Mittelwertzentrum eines Segments projizierte einerseits erregend in die Neuronen der Klasse 1 des gleichen Segments, die sich jeweils sowohl im sensorischen als auch im motorischen Zentrum befanden. Weiterhin projizierte es unter Zwischenschaltung der afferent projizierenden Neuronen der Klasse 4 wiederum erregend in die Neuronen der Klasse 1 in allen kopfseitig gelegenen Segmenten. Auch diese Neuronen befanden sich jeweils im motorischen und im sensorischen Zentrum eines jeden kopfseitigen Segments.
So erreichte die zur Voraktivierung genutzte Mittelwerterregung jedes Mittelwertkerns alle Segmente, die auf gleicher oder größerer Höhe lagen.
Und durch die laterale Hemmung, verursacht durch die Interneuronen in den sensorischen und motorischen Zentren, standen alle gleichartigen Mittelwertsysteme in gegenseitiger Konkurrenz, sowohl bezüglich der Segmente einer Körperhälfte als auch bezüglich beider Körperhälften.
Denn wir unterstellen, dass die laterale Hemmung zunächst auf Signale beschränkt war, die signalverwandt waren, also z. B. der gleichen Rezeptorenart stammten bzw. aus gleichartigen Mittelwertzentren stammten.
Ebenso projizierten in jedem Segment die Neuronen der Klasse 6 nicht nur in die Mittelwertzentren des gleichen Segments, sondern auch in alle schwanzseitig gelegenen Segmente.
Die neuronale Konkurrenz der Signale von verschiedenen Segmenten einer Körperhälfte sollte gravierende Folgen für den Aufbau des Körpers der segmentierten Tiere nach sich ziehen. Diese werden sichtbar durch eine zunehmende Spezialisierung, in deren Folge sich zwischen den Segmenten eine Arbeitsteilung entwickelt, so dass nicht mehr jedes Segment für alles zuständig ist.
Theorem der neuronalen Organisation der segmentierten Bilateria
In segmentierten Bilateria, deren Linie zu den Wirbeltieren führte, besaß anfänglich jedes Segment die gleiche neuronale Architektur. Jedes Segment besaß ein linkes und ein rechtes sensorisches Zentrum, ein linkes und ein rechtes motorisches Zentrum sowie verschiedene Mittelwertzentrum, ebenfalls jeweils bilateral links und rechts angeordnet.
In jedem Segment mit Ausnahme des Kopfsegments gab es auf jeder Körperseite eine afferente Projektion vom sensorischen Zentrum des Segments zum sensorischen Zentrum des darüber liegenden Segments. Diese Projektion wurde von Neuronen der Klasse 4 realisiert, deren Axone die Längsverbindungen bildeten, die als sensorische Konnektive bezeichnet werden. Die Axone dieser Neuronen haben in jedem Segment eine konstante Länge und reichen genau bis zum sensorischen Zentrum des darüber angeordneten Segments.
In jedem Segment mit Ausnahme des Schwanzsegments gab es eine efferente Projektion vom motorischen Zentrum eines Segments zum motorischen Zentrum des darunter befindlichen Segments. Diese Projektion wurde von Neuronen der Klasse 5 bewirkt, deren Axone ebenfalls Längsverbindungen bildeten, die als motorische Konnektive bezeichnet werden. Die Axone dieser Neuronen haben in jedem Segment eine konstante Länge und reichen genau bis zum motorischen Zentrum des darunter angeordneten Segments.
Jedes sensorische Zentrum empfing die Erregung der ipsilateralen Rezeptoren des gleichen Segments (Eigensignale) sowie über Neuronen der Klasse 4 die Erregung der ipsilateralen Rezeptoren aller tieferliegenden, schwanzseitigen Segmente (Fremdsignale). Zusätzlich empfingen die Neuronen der Klasse 1 die Vorerregung aus den ipsilateralen Mittelwertzentren des gleichen Segments sowie über Neuronen der Klasse 4 die ipsilaterale Mittelwerterregung aller tieferliegenden, schwanzseitigen Segmente. Die Neuronen der Klasse 1 übertrugen diese Vorerregung auf die Neuronen der Klasse 3, ebenso auf die Neuronen der Klasse 4.
Die Axone der Neuronen der Klasse 3 bildeten die Querverbindungen, die als Kommissuren bezeichnet werden, sie endeten für die Eigensignale des Segments an den ipsilateralen Motoneuronen dieses Segments. Fremdsignale erreichen über die Axone der Neuronen der Klasse 3 die Neuronen der Klasse 5 im ipsilateralen motorischen Zentrum und werden absteigend zu den Zielsegmenten weitergereicht.
Jedes motorische Zentrum projizierte über Neuronen der Klasse 2 (Kreuzkommissuren) erregend in hemmende Interneuronen des motorischen Zentrums der Gegenseite des gleichen Segmentes und bewirkte dort die kontralaterale motorische Hemmung auf Segmentebene.
Auf die gleiche Art projizierte jedes sensorische Zentrum über Neuronen der Klasse 2 (Kreuzkommissuren) erregend in hemmende Interneuronen des sensorischen Zentrums der Gegenseite des gleichen Segmentes und bewirkte dort die kontralaterale sensorische Hemmung auf Segmentebene.
Ebenso erreichte die Erregung des motorischen Zentrums über Neuronen der Klasse 6 die gleichseitigen Mittelwertzentren des Segments sowie, falls vorhanden, die entsprechenden Mittelwertzentren der darunter gelegenen Segmente.
Die Erregung des sensorischen Zentrums erreichte ebenso über Neuronen der Klasse 6 die gleichseitigen Mittelwertzentren des Segments sowie, falls vorhanden, die entsprechenden Mittelwertzentren der darunter gelegenen Segmente.
Jedes Mittelwertzentrum einer Körperseite projizierte in die Neuronen der Klasse 1 des gleichseitigen sensorischen und motorischen Zentrums der gleichen Körperseite, wodurch diese eine mittelwertabhängige Vorerregung erhielten, sowie über Neuronen der Klasse 4 in Neuronen der Klasse 1 der ipsilateralen sensorischen und motorischen Zentren aller kopfseitig darüber befindlichen Segmente.
Sowohl im sensorischen als auch im motorischen Zentrum jeder Körperseite und jedes Segments gab es hemmende Interneuronen, die eine laterale Hemmung verursachten und zur Kontrastverstärkung stärkerer Signale und zur Abschwächung schwächerer Signale führten. Gleiches galt für alle Mittelwertkerne.
Das Nervensystem der segmentierten Bilateria, deren Linie zu den Wirbeltieren führte, bestand aus zwei Strickleitersystemen, je einem für jede Körperhälfte. Die sensorischen Zentren entsprachen den Ganglien des sensorischen Leiterholms, die motorischen Zentren den Ganglien des motorischen Leiterholms. Die sensorischen Ganglien waren mit den motorischen Ganglien der gleichen Körperseite über Kommissuren verbunden.
Die sensorischen und motorischen Ganglien waren mit denen der Gegenseite über Kreuzkommissuren verbunden, die der kontralateralen Hemmung dienten.
Die Axone der verschiedenen Neuronenklassen verliefen angenähert in rechten Winkeln zueinander, dies gilt auch für die Kreuzkommissur.
In späterer Evolutionszeit konnten die Axone der Neuronen der Klasse 5 und 6 ihre Länge, die ursprünglich genau bis zum nächsten Segment reichte, vervielfachen, so dass ihre Axonlänge sich (später) auch über mehrere Segmente erstrecken konnte.
Für das Verständnis der topologischen Organisation ist es erforderlich, sich Gedanken zu machen über die Wachstumsrichtungen der Axone der verschiedenen Neuronenklassen, die mit hoher Wahrscheinlichkeit durch die Konzentrationsgradienten von neuronalen Markersubstanzen gesteuert werden.
Richtungstheorem des Strickleiter-Nervensystems
In einem idealisierten Strickleiter-Nervensystem bilden die Axone der Konnektivneuronen mit den Axonen der Kommissurneuronen rechte Winkel und spannen eine Ebene auf, zu der die Axone der Mittelwertneuronen ebenfalls senkrecht ausgerichtet sind. Die Dendriten orientieren sich in etwa entgegengesetzt zu den Axonen. Räumliche Deformationen des Körpers können systematische topologische Abweichungen von den Idealrichtungen bewirken.
Die verschiedenen Projektionsneuronen leiten auf ihren Axonen Signale weiter.
Da wir den Projektionsneuronen im Nervensystem Neuronenklassen zuordnen können, ist es möglich, auch die Signale in Signalklassen zu unterteilen. Hierbei sei die Signalklasse ableitbar aus der Neuronenklasse des Neurons, welches dieses Signal auf seinen Axonen transportiert. Daher gibt es auch sechs Signalklassen.
Dann sind beispielsweise folgende Aussagen gleichwertig:
- Die Neuronen der Klasse 6 projizieren ins Mittelwertzentrum.
- Die Signalklasse 6 erreicht das Mittelwertzentrum.
Wir stellen uns das Strickleiter-Nervensystem in einer Anordnung vor, bei der sich oben der Kopf und unten der Schwanz des Lebewesens befindet. Günstig war ein gleichmäßiger, kreuzungsfreier Verlauf der Axone, möglichst getrennt nach den verschiedenen Neuronenklassen.
Wie in der nachfolgenden Abbildung symbolisch dargestellt, befinden sich die Konnektivneuronen der Klassen 2 und 3 jeweils innen neben den zugehörigen Kommissurneuronen der Klassen 4 und 5, weil sie die waagerecht verlaufende Verbindung herstellen und sich so an der Position befinden, die kurze Leitungswege sichert.
Die nach oben und unten projizierenden Kommissurneuronen der Klasse 4 und 5 befinden sich dagegen außen, so dass ebenfalls ein kreuzungsfreier Verlauf ihrer Axone möglich ist.
Die Neuronen der Klasse 6, die zu den Mittelwertzentren projizieren, befinden sich auf der motorischen Seite ganz außen, weil ihre Ziele sich außerhalb des Strickleiter-Nervensystems befinden. Die Lage der Neuronen der Klasse 1 im Inneren der sensorischen Seite ist hypothetisch.
Die langen Axone der kopfwärts projizierenden Neuronen der Klasse 4 verliefen im sensorischen Teil des Strickleitersystems an dessen Außenseite. Gleiches galt für die schwanzwärts projizierenden Axone der Neuronen der Klasse 5. Diese zwei Klassen von Axonen waren besonders lang, jedes von ihnen musste jeweils eine Segmentlänge des Körpers überbrücken. Daher waren sie zunächst recht dick, damit die Signaldämpfung die Aktionspotentiale nicht zum Erliegen brachte. Gleichzeitig dienten diese größeren Axondurchmesser der Erreichung einer höheren Ausbreitungsgeschwindigkeit der Aktionspotentiale. Bei einfachen Bilateria gibt es noch heute diese dicken Axonbündel, die vom Kopf zum Schwanz verlaufen und teils als Riesenfasern bezeichnet werden. Später bildete sich Myelin als elektrische Abschirmung der Axone der Konnektivneuronen der Klassen 4 und 5. Sie sind im Rückenmark der Wirbeltiere ebenfalls außen angeordnet und bilden dort die weiße Substanz.
Die Axone der Kommissurneuronen der Klasse 2 und 3 dagegen verliefen im Innenteil des Strickleitersystems, sie zogen jeweils waagerecht zu den Zielneuronen der Klassen 4 und 5. Wegen ihrer Kürze brauchte sich hier keine Myelinschicht auszubilden, diese Axone gehörten damit zur grauen Substanz.
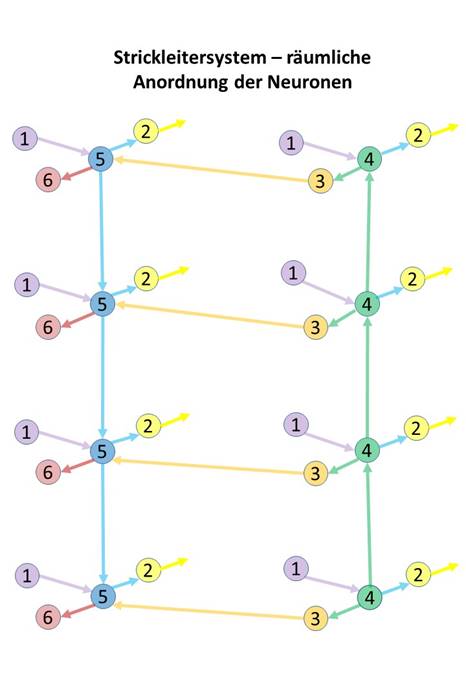
Abbildung 2- Räumliche Anordnung der Neuronenklassen
In Abbildung 2 ist nur eine Strickleiter des tetraneuralen Nervensystems dargestellt. Das zweite Strickleitersystem der anderen Körperhälfte wird über die Neuronen der Klasse 2 mit diesem verbunden, wobei die Kreuzkommissuren der Neuronen der Klasse 2 in beide Richtungen verlaufen. Nicht dargestellt sind die Mittelwertsysteme.
Theorem der kontralateralen Hemmung der neuronalen Zentren auf Segmentebene
Jedes neuronale Zentrum (sensorisches sowie motorisches Zentrum, Mittelwertzentrum) in jedem Segment und auf jeder Körperseite projiziert erregend in hemmende Interneuronen des gleichen neuronalen Zentrums der gegenüberliegenden Körperseite. Beide stehen in Konkurrenz zueinander.
Die Projektion zur kontralateralen Seite des gleichen Segments erfolgte über eingeschaltete Zwischenneuronen, die sich etwa an der (gedachten) Trennebene der beiden Körperhälften befanden. Ein Grund war der Wechsel des Markers, der die Körperseite kennzeichnete. Jede Körperhälfte besaß wahrscheinlich einen eigenen Körperseitenmarker. Beim Übergang von einer Körperhälfte zur anderen wechselte der Körperseitenmarker, so dass an der Grenzstelle ein Zwischenneuron nötig wurde, welches nun auf den kontralateralen Seitenmarker ansprach und dessen Axon zum entsprechenden neuronalen Zentrum führte.
Die Zwischenneuronen, die ebenfalls Projektionsneuronen waren, bildeten im Verlaufe der Evolution eigene Kerne, die wir als Seitenwechselkerne bezeichnen. Es gab pro Etage auf jeder Körperseite je einen sensorischen und einen motorischen Seitenwechselkern.
Theorem der Seitenwechselkerne
In jedem Segment bildeten sich auf jeder Körperseite je ein sensorischer und ein motorischer Seitenwechselkern. Der sensorische empfing den Output des sensorischen Kerns dieser Körperseite und projizierte erregend in den sensorischen Kern der Gegenseite, wo er an hemmenden Interneuronen endete und die kontralaterale Hemmung bewirkte. Der motorische empfing den Output des motorischen Kerns dieser Körperseite und projizierte erregend in den motorischen Kern der Gegenseite, wo er an hemmenden Interneuronen endete und ebenfalls die kontralaterale Hemmung bewirkte.
Die Seitenwechselkerne befanden sich in der Nähe der Trennebene zwischen beiden Körperhälften. Der Input erreichte die Seitenwechselkerne über Neuronen der Klasse 2. Wir unterstellen, dass die Neuronen im Seitenwechselkern ebenfalls zur Neuronenklasse 2 gehören, jedoch auf den kontralateralen Seitenmarker ansprachen.
Ob die kontralaterale Hemmung der Mittelwertkerne ebenfalls über solche Seitenwechselkerne erfolgte, soll hier angesichts der geringen Neuronenanzahl dieser Kerne nicht weiter ausdiskutiert werden.
Monografie von Dr. rer. nat. Andreas Heinrich Malczan