Gehirntheorie der Wirbeltiere
ISBN
978-3-00-064888-5
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
3.5 Die topologische Wohlordnung der Neuronen im Startsystem
Die tierischen Lebewesen brachten im Verlauf der Evolution eine ungeheure Vielfalt an Rezeptoren hervor, mit denen unterschiedlichste Umweltreize in neuronale Signale transformiert werden können.
War die Anzahl der verschiedenen Rezeptorenarten anfangs nur gering, und gab es zu jeder von ihnen zunächst nur wenige Rezeptoren, so konnte ein doppeltes Strickleitersystem in Form eines tetraneuralen Nervensystems die Signale bei den segmentierten Bilateria recht gut transportieren. Wenn nun aber die Anzahl der Rezeptorenarten anstieg, und zusätzlich auch die Anzahl der Rezeptoren gewaltig zunahm, so konnte ein einzelner Neuronenstrang die riesige Menge von Axonen gar nicht aufnehmen. Der Strang, nehmen wir hier einmal den afferent projizierenden sensorischen Strang einer Körperseite, wurde zunehmend zu einer Art Flachbandkabel aus Axonen, die in einer dicken Schicht angeordnet waren. Die vier Nervenstränge des tetraneuralen Systems, zwei sensorische und zwei motorische, bildeten eine Art Rohr, dessen Inneres funktionslos war und einen Hohlraum bildete. Dieser wurde mit einer Flüssigkeit gefüllt, die inkompressibel war und dem hohen Wasserdruck auch in größerer Tiefe widerstehen konnte. So bildete sich das Neuralrohr der künftigen Wirbeltiere.
Theorem der Neuralrohrbildung
Das bilateral symmetrische Neuralrohr entwickelte sich aus dem tetraneuralen Nervensystem durch eine im Verlaufe der Evolution sehr starke Zunahme sowohl der Rezeptoren einer Rezeptorenart als auch der Anzahl der verschiedenen Rezeptorenarten. Die linke und die rechte Hälfte des Neuralrohrs besteht jeweils aus der Parallelschaltung vieler einzelner Strickleitersysteme, die sensorischen und motorischen Zentren der Segmente sind in der bereits beschriebenen Weise miteinander verschaltet. Alle Strukturelemente des tetraneuralen Nervensystems sind analog im Neuralrohr realisiert, etwa die Mittelwertkerne. Ebenso wird die kontralaterale Hemmung verwirklicht. Das Neuralrohr und das tetraneurale Nervensystem derjenigen segmentierten Bilateria, deren Linie zu den Wirbeltieren führt, unterscheiden sich (anfänglich) nur durch die viel größere Anzahl der Neuronen. Die Ursache dafür ist die enorme Zunahme der Anzahl der Rezeptoren sowohl der gleichen Modalität als auch verschiedener Modalitäten.
Die sensorischen Konnektive bilden im Neuralrohr das Hinterhorn, die motorischen das Vorderhorn.
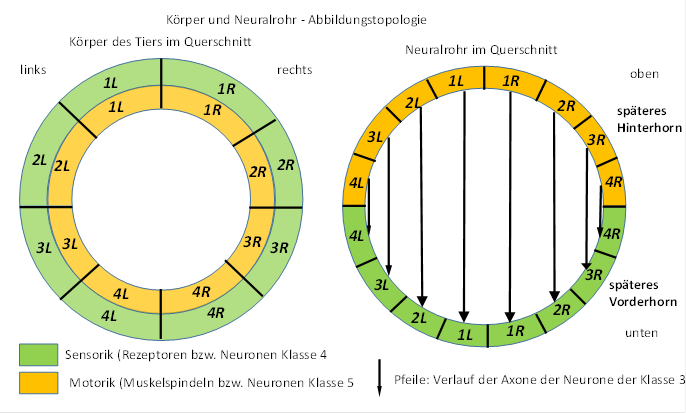
Abbildung 6- Körper und Neuralrohr - Abbildungstopologie
Richtungstheorem des Neuralrohr-Nervensystems
In einem idealisierten Neuralrohr-Nervensystem bilden die Axone der Konnektivneuronen mit den Axonen der Kommissurneuronen rechte Winkel. Räumliche Deformationen des Körpers können zu topologischen Abweichungen von den Idealrichtungen bewirken.
Die Ergebnisse des amerikanischen Connectome-Projektes, bei dem der Axonverlauf im menschlichen Gehirn durch computertomografische Untersuchungen der Diffusionsgradienten von Wassermolekülen untersucht wird, bestätigen diese räumliche Orientierung. Axone gleicher Orientierung verlaufen gleichförmig nebeneinander, ohne sich zu vermischen. Abzweigende Axone bilden häufig rechte Winkel zum bisherigen Axonverlauf.
Klassifizierungstheorem für Neuronen
Das Richtungstheorem erlaubt es, die Projektionsneuronen nur auf Grund ihres Axonverlaufs den sechs verschiedenen Neuronenklassen zuzuordnen. Neuronen, deren Axone kopfwärts (nach oben) verlaufen, gehören zur Neuronenklasse 4. Schwanzwärts (nach unten) verlaufende Axone treten bei Neuronen der Klasse 5 auf. Waagerecht verlaufende Axone gehören zu den Neuronenklassen 3 und 2, wobei die Axone der Neuronen der Klasse 3 motorische Ziele ansteuern, während die Axone der Klasse 2
immer zur Gegenseite in analoge neuronale Zentren projizieren. Axone der Neuronen der Klasse 6 ziehen zu Mittelwertzentren, während die Neuronen der Mittelwertzentren ihre Axone zu den Neuronen der Klasse 1 aussenden.
In gewisser Weise könnten Axone der Neuronen der Klassen 4 und 5 mit den Axonen der Klassen 6 und 1 verwechselt werden, wenn man sie nur in kleinen Gebieten betrachtet, ohne ihre Endigungsneuronen zu kennen. Diese Mehrdeutigkeit entfällt, sobald man die Ursprungs- und die Zielstrukturen erkennt.
Würde man das Neuralrohr entlang seiner Längsachse durchschneiden, wobei der Schnitt etwas seitlich orientiert ist und sowohl das Hinterhorn als auch das Vorderhorn einer Körperseite durchschneidet, so erhielte man im Schnittbild wieder das urtümliche Strickleitersystem. Natürlich geht durch die enorme Anzahl der Neuronen der Eindruck der ursprünglichen Segmentierung verloren, da die Neuronen eines sensorischen oder motorischen Zentrums bei Platzmangel nach oben und unten ausweichen, wo bereits die Neuronen der Nachbarsegmente ebenso ihren Platz zu behaupten versuchen. So entsteht eine Fläche, auf die sich die ursprünglichen sensorischen und motorischen Zentren verteilen.
Die ursprüngliche Segmentierung ist jedoch noch gut nachzuvollziehen anhand der Spinalganglien, die als eigenständige Axonbündel in das Neuralrohr treten.
Wir unterteilen das Neuralrohr in einen Rumpfanteil und einen Kopfanteil. Wir stellen uns nun eine beliebige Schnittebene vor, die den Körper des Tieres in zwei Teile teilt. Die Schnittrichtung möge rechtwinklig zur Körperachse verlaufen. Im Schnittbild - welches etwa kreisförmig aussieht - untersuchen wir die Anordnung der Axone der aufwärts bzw. abwärts projizierenden Konnektive.
Wir beschränken uns zunächst auf die Axone einer Modalität. Ihre Signale entstammen einer ausgewählten Rezeptorenart, beispielsweise den Muskelspindeln. Diese bildeten sich relativ spät heraus, davor übernahmen freie Nervenendigungen die Messung der Muskelspannung.
Theorem der Modalitätenringe und Segmentringe der Axone im Rumpfteil des Neuralrohrs
Die Axone der aufwärts projizierenden Neuronen der Klasse 4 sowie die Axone der abwärts projizierenden Neuronen der Klasse 5 einer jeden Modalität bilden im Neuralrohr einen Modalitätenring. Innerhalb eines jeden Modalitätenrings befinden sich so viele Segmentringe, wie es Segmente mit den Rezeptoren dieser Modalität gibt.
Die Segmentringe einer Modalität sind ineinander geschachtelt und wohlgeordnet. Innen befindet sich der Segmentring mit der höchsten Segmentnummer, wobei die Nummerierung am Kopfsegment beginnt. Nach außen nimmt die Segmentnummer stetig ab. Der äußerste Segmentring beinhaltet die Axone, deren Rezeptoren im höchsten kopfseitigen Segment zu finden sind.
Im Schnittbild ähnelt das Neuralrohr einem Baumstamm mit Jahresringen, allerdings sind hier Gruppen von nebeneinanderliegenden Jahresringen zu Modalitätenringen zusammengefasst.
Die Ursache für diese Wohlordnung im Neuralrohr liegt darin begründet, dass die erste Rezeptorenart, die hervorgebracht wurde, für die kopfseitige Projektion die Neuronen der Klasse 4 kontaktierte. Deren Axone wuchsen einfach gerade nach oben. Jedes höhere Segment lagerte seine sensorischen Projektionsaxone außen an die bereits bestehenden an. So ergab jedes Segment einen Segmentring von aufsteigenden Axonen. Im motorischen Teil des Neuralrohrs war es analog. Hier befanden sich die absteigenden Axone zum untersten Schwanzsegment innen und bildeten den innersten Segmentring dieser Modalität. Die Axone zum zweituntersten Segment lagerten sich den beschriebenen außen an und bildeten den nächsten Segmentring. Dies ging so weiter, bis das erste Kopfsegment erreicht wurde. Die Axone seiner abwärts projizierenden Neuronen der Klasse 5 lagerten sich beim bestehenden Segmentringsystem ebenfalls außen an und bildeten den äußeren Segmentring. Die Gesamtheit aller Segmentringe der ersten Modalität bildete so im Schnittbild auf der Höhe des ersten Segmentes einen Modalitätenring.
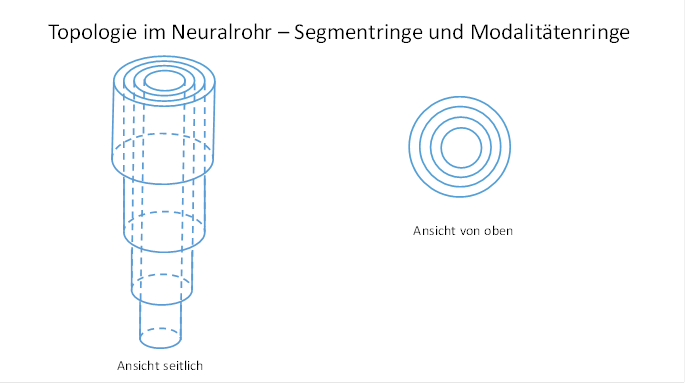
Abbildung 7- Topologie im Neuralrohr - Segmentringe und Modalitätenringe
Als sich eine zweite Rezeptorenart bildete, wurden ihre Projektionsaxone außen an den ersten Modalitätenring angelagert. Dies erfolgte wiederum auf die beschriebene Weise in Segmentringen, die an die Höhe des Segments gekoppelt waren.
Die Modalitätenringe der verschiedenen Modalitäten lassen einen Schluss zu auf die Reihenfolge ihrer evolutionären Herausbildung zu. Die Modalitätenringe der früher herausgebildeten Rezeptorenarten lagen innen, die der evolutionär später entwickelten dagegen außen.
Jeder Segmentring bestand aus vier Viertelringen, denn im ursprünglichen tetraneuralen Nervensystem gab es zwei sensorische und zwei motorische Axonstränge, die die Konnektive bildeten und jeweils aufwärts bzw. abwärts projizierten.
Nachdem wir den Axonverlauf und die Wohlordnung der Axone beschrieben haben, wenden wir uns der räumlichen Anordnung der beteiligten Neuronen zu, die diese Axone besitzen.
Theorem der segmentierten Körperabbildungen im Rumpfbereich
Die sensorischen und die motorischen Konnektivneuronen der verschiedenen Modalitäten bilden in jedem Segment eine annähernd zylindrische Mantelfläche, die ein topologisches Körperabbild der signalliefernden Rezeptoren der Segmente darstellt. Hierbei entsteht auf der sensorischen Seite ein sensorisches Körpermodell auf der Mantelfläche eines Halbzylinders, während auf der motorischen Seite ein analoges motorisches Körpermodell entsteht. Nachbarschaftsbeziehungen der Rezeptoren bleiben im sensorischen Körperabbild erhalten. Nachbarschaftsbeziehungen der Zielmuskeln bleiben im motorischen Körperabbild erhalten.
Wir fassen den sensorischen und den motorischen Teil des Körpermodells einer Modalität zu einem Modalitätenzylinder zusammen, auf dessen Mantelfläche sich die sensorische und die motorische Körperabbildung befinden. Die Modalitätenzylinder der verschiedenen Modalitäten sind ineinander geschachtelt, im Inneren verläuft der Spinalkanal des Neuralrohrs. Innen befinden sich die evolutionär älteren, weiter außen die evolutionär jüngeren Modalitäten. Die Trennebene zwischen dem sensorischen und dem motorischen Teil des Neuralrohrs ist die Symmetrieebene, jede orthogonal auf ihr stehende Gerade verbindet gleiche Körperpunkte der beiden Abbildungen. Alle Axone der Neuronen der Klasse 3 verlaufen ebenfalls orthogonal zu dieser Trennebene.
In jedem Segment befindet sich ein derartiges Gebilde aus ineinander geschachtelten Modalitätenzylindern. Da diese ihren Input von unten (schwanzseitig) bekommen, können in den Körpermodellen auch nur die unten befindlichen Segmente erfasst sein. Die Kopfsegmente enthalten dagegen für jede Modalität eine Körperabbildung aller Rumpfsegmente. Nur dort kann eine Signalverarbeitung die Signale des gesamten Körpers mit einbeziehen. Daher muss sich das Gehirn kopfseitig befinden.
Bei den Rumpfmodalitäten werden in das sensorische Körperabbild im Segment mit der Nummer k (vom Kopf aus gezählt) nur die signalliefernden Rezeptoren einbezogen, also nur Segmente mit den Nummern k bis n, wenn n die Segmentanzahl darstellt. Die oberhalb des betreffenden Segments befindlichen Rezeptoren beliefern (normalerweise) nur die höheren Segmente.
An dieser Stelle ist es angebracht, sich Gedanken über die Topologie der Kerne des siebenten Segments zu machen, welches das Eingangs- und Ausgangssegment des Urhirns war. Es enthielt wichtige Projektionskerne, so etwa den Nucleus ruber, den Nucleus olivaris und den Nucleus Purkinje, aus dem sich das spätere Cerebellum entwickeln sollte.
Theorem der motorischen Körperabbildungen im Nucleus ruber, Nucleus olivaris und Nucleus Purkinje
Die motorischen Konnektivneuronen der Klasse 5 einer Körperhälfte bilden im siebenten Segment die Mantelfläche eines Halbzylinders, die ein topologisches Körperabbild aller ipsilateralen Motoneuronen des Lebewesens darstellt, welches nach Segmentringen geordnet war. Nachbarschaftsbeziehungen zwischen den Muskeln und den Motoneuronen bleiben dabei erhalten. Neben dieser Neuronenfläche existiert ebenso eine zweite Mantelfläche, die von den Kommissurneuronen der Klasse 2 gebildet wird, die den Anfang der Neuronenkette für die kontralaterale Hemmung darstellen, die in einer Punkt-zu-Punkt-Verbindung über den Nucleus olivaris zum Purkinjekern und von dort zum kontralateralen Nucleus ruber verläuft.
Die Axone der Neuronenschicht der Klasse 2 des Nucleus ruber enden im Nucleus olivaris.
Da sie unvermischt verlaufen, bilden die Neuronen des Nucleus olivaris ebenfalls ein nach Segmentringen wohlgeordnetes Körperabbild der Motoneuronen der ipsilateralen Körperhälfte. Diese Topologie übertragen die Axone der Projektionsneuronen auf den Nucleus Purkinje.
Anfänglich sind der Nucleus Purkinje und der Nucleus ruber direkt benachbart, später trennen sich die GABAergen Neuronen des Nucleus Purkinje ab und bilden einen eigenständigen Kern. Die Projektionsaxone seiner Kommissurneuronen verlaufen waagerecht wohlgeordnet zum Nucleus ruber, ohne sich gegenseitig zu kreuzen.
Der Nucleus Purkinje projizierte nach dem gleichen Schema in einer Punkt-zu-Punkt-Abbildung hemmend in den kontralateralen Nucleus ruber und bewirkte dort die kontralaterale Hemmung der zugeordneten motorischen Gegenspieler. Hierbei dockt jedes GABAerge Axon an dem Projektionsneuron der Klasse 5 im Nucleus ruber an, welches zum motorischen Gegenspieler projiziert. Dadurch wird die kontralaterale Hemmung realisiert.
Durch die starke Zunahme der beteiligten Neuronen wurden alle diese ein- bzw. zweischichtigen Halbzylinderflächen, die eigentlich Neuronenschichten darstellen, im Verlauf der weiteren Evolution kontinuierlich verformt, zusammengedrängt, teils in Falten gelegt (um Platz zu sparen). Sie erhielten dadurch ein sachartiges Aussehen. Das Abbildungsprinzip blieb jedoch prinzipiell erhalten. Jeder Kern bildete eine nach Segmentringen wohlgeordnete Körperabbildung. Der Nucleus ruber, der Nucleus olivaris und der Nucleus Purkinje stellten nach Körpersegmenten wohlgeordnete Körperabbildungen der Motoneuronen einer Körperhälfte dar, wobei generell eine Punkt-zu-Punkt-Verbindung eingehalten wurde.
Die Bilateralität blieb auch hier erhalten, jeder der Kerne war ipsi- und kontralateral vertreten. So blieb es nicht aus, dass sich die Kreuzkommissuren real kreuzten, auch wenn die Axone ansonsten unvermischt verliefen.
Günstig war die Anordnung in halbzylindrisch angeordneten Strukturen, so dass genügend Platz vorhanden war, die waagerecht verlaufenden Projektionsaxone in Richtung des kontralateralen Nucleus ruber kreuzungsfrei und unvermischt von einem Kern zum nächsten verlaufen zu lassen, bis der Zielkern auf der Gegenseite erreicht war, wobei sie natürlich den Axonen der Gegenseite begegnen mussten. Die beteiligten Axone der Kommissurneuronen der Klassen 2 verliefen ungestört waagerecht, während die Axone der Projektionsneurone der Klassen 4 und 5 senkrecht nach oben bzw. unten verliefen, ohne sich gegenseitig zu stören (wenn der Kopf sich oben befand).
Bei der Betrachtung der topologischen Wohlordnung im Startsystem wurden die Neuronenklassen 1 und 6 bewusst weggelassen, um das Modell nicht unnötig zu verkomplizieren. Im Nachhinein ist es nicht schwierig, diese Neuronenklassen in das Modell einzufügen, ebenso die hemmenden Interneuronen, die der lateralen Nachbarhemmung dienten.
Im Verlauf der Evolution sollten weitere Signale den Weg in den Olivenkern finden, die teils auch nichtmotorischer Art waren. Die Punkt-zu-Punkt-Verbindung war nur anfangs gegeben und ging spätestens in der zweiten Erweiterungsphase des sich bildenden Cerebellums verloren.
- ← 3.4 Die Grundstrukturen im Urhirn der segmentierten Bilateria
- 3.6 Die topologische Wohlordnung im Kopfbereich →
Monografie von Dr. rer. nat. Andreas Heinrich Malczan