Gehirntheorie der Wirbeltiere
ISBN
978-3-00-064888-5
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
4.3 Die zweite Erweiterungsphase des urtümlichen Pontocerebellums
Mit dem starken Anwachsen der Rezeptorenanzahl im Verlaufe der Evolution wurde es vorteilhafter, die cortikalen Mittelwerterregungen zusammenzufassen, bevor sie einer Auswertung zugeführt wurde. Daher konvergierten mehrere cortikale Mittelwertneuronen der Klasse 6 auf ein gemeinsames Mittelwertneuron des Nucleus subthalamicus. Dieser stellte den Mittelwertkern der thalamischen Etage dar.
Ursprüngliche projizierte jeder Cortexcluster über sein Mittelwertneuron in den Nucleus subthalamicus. Nun jedoch projizierte eine Gruppe von benachbarten Cortexclustern in ein solches Neuron.
Denkt man sich das anfänglich wurmförmige Lebewesen senkrecht angeordnet, so dass sich oben der Kopf und unten der Schwanz befindet, so hatte der Cortex als Fläche eine ähnliche, allerdings stark verkleinerte Form. Diese war zudem in Modalitätenstreifen und Segmentstreifen unterteilt. Der Nucleus subthalamicus als Körpermodell der cortikalen Mittelwerte besaß eine analoge Anordnung. Bei der Mittelwertbildung war es vorteilhaft, die Modalitätengrenzen und die Segmentgrenzen einzuhalten. Daher wurden nur Cluster zu einem gemeinsamen Mittelwert zusammengefasst, die waagerecht nebeneinander eine Clustergruppe zu einer Modalität bildeten.
Definition: Konvergenzgrad des Nucleus subthalamicus
Wenn die Mittelwertneuronen einer Clustergruppe aus n nebeneinander angeordneten Cortexclustern in ein gemeinsames Outputneuron des Nucleus subthalamicus projizieren, dann bezeichnen wir die Zahl n als den Konvergenzgrad.
Der Konvergenzgrad ist also die Anzahl der Cortexcluster, die in ein gemeinsames Outputneuron des Nucleus subthalamicus projizieren.
Nach der Konvergenz der cortikalen Mittelwertsignale auf die Neuronen des Nucleus subthalamicus war die Cerebellumrinde des Pontocerebellums bezüglich der Körnerzellen immer noch ein Cortexmodell. Doch die Anzahl der Outputneuronen des Nucleus subthalamicus, die in den Nucleus olivaris projizierten, hatte sich - bezogen auf die Anzahl der Cortecluster - deutlich verringert.
Theorem der Clustergruppen im Nucleus subthalamicus
Jede Clustergruppe, bestehend aus benachbarten Cortexclustern, wird im Nucleus subthalamicus durch ein gemeinsames Mittelwertneuron vertreten.
Theorem vom Nucleus subthalamicus als Mittelwert-Körpermodell
Der Nucleus subthalamicus stellt ein vergröbertes Körpermodell der Cortexrinde dar, bei dem jede cortikale Clustergruppe durch ein Mittelwertneuron vertreten wird.
Die Konvergenz der Mittelwertsignale im Nucleus subthalamicus war sicherlich sinnvoll. Da jeder Mittelwertkern mit den Mittelwertkernen anderer Segmente Daten austauschte, war eine Reduktion der signalführenden Axone materialsparend. Fiel ein Neuron aus, so war die Mittelwertbildung nur geringfügig gestört, da die übrigen Axone ebenfalls Mittelwerte lieferten. Der Konvergenzgrad durfte andererseits nicht zu groß werden, weil sonst die Zuordnung eines Mittelwertes zur signalerzeugenden Stelle des Körpers nicht mehr präzise genug war.
Wir postulieren hier, dass das Mittelwertmodell des Nucleus subthalamicus die Segmentstruktur des Körpers widerspiegelte. Daher wurden von den eintreffenden Axonen jeweils nur diejenigen in einem gemeinsamen Outputneuron zusammengefasst, die Signale aus dem gleichen Körpersegment repräsentierten. Damit verliefen die Dendriten der Outputneuronen in etwa parallel zueinander nach links und rechts, wenn sich im Körpermodell der Kopf oben und der Schwanz unten befand. Diese Projektion berücksichtigte auch die Modalitäten, die streng auseinandergehalten wurden.
Diese Signalkonvergenz wurde im Nucleus olivaris durch eine entgegen gerichtete Signaldivergenz wieder rückgängig gemacht. Denn parallel zur Signalkonvergenz im Nucleus subthalamicus trat eine Signaldivergenz im Nucleus olivaris auf. Hier verursachte der Ausfall eines einzelnen Neurons im Nucleus olivaris eine empfindliche Störung der Motorik. Daher fand im Nucleus olivaris eine zunehmend stärkere Signaldivergenz statt. Es wurden daher je zwei Inputsignale auf n Outputsignale verteilt. Dieses Prinzip wurde auch in den Mittelwertgebieten des Nucleus olivaris angewendet, die ins Pontocerebellum projizierten.
Wenn nun zwei benachbarte Clustergruppen C1 und C2 (des gleichen Segments) in den Nucleus subthalamicus projizierten und das Striosomensystem daraus zwei Kletterfasersignale fA und fB für das Pontocerebellum generierte, so trafen diese bei zwei benachbarten Inputneuronen des Nucleus olivaris ein.
Wir gehen davon aus, dass diese Inputsignale im Nucleus olivaris divergent auf eine Gruppe von Outputneuronen verteilt wurden, die in einer Zeile angeordnet waren. Mit fortschreitender Evolution nahm ihre Anzahl zu.
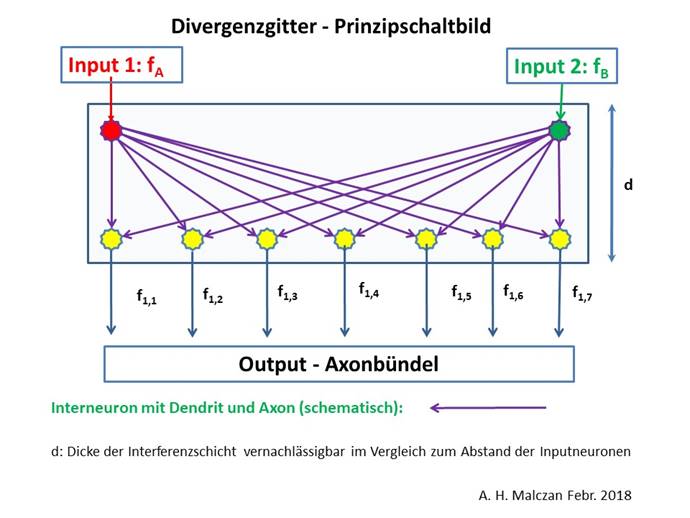
Abbildung 37- Signaldivergenz im Nucleus olivaris
Wir wiederholen hier eine frühere Definition sinngemäß:
Definition: Divergenzgrad im Nucleus olivaris
Werden im Nucleus olivaris zwei Inputsignale fA und fB durch Signaldivergenz auf eine Zeile von n Outputneuronen verteilt, so bezeichnen wir die Zahl n als Divergenzgrad des Nucleus olivaris.
Wir unterstellen nun, dass im Verlaufe der Evolution der Konvergenzgrad im Nucleus dentatus und der Divergenzgrad im Nucleus olivaris in etwa gleich groß waren.
Theorem der Umkehrung der Signalkonvergenz im Nucleus dentatus
Im Nucleus dentatus wurde die Signalkonvergenz des Nucleus olivaris durch eine annähernd gleich große Signaldivergenz rückgängig gemacht.
Die Verteilung der Kletterfasersignale erfolge durch parallele Divergenz. Die abgeleiteten Kletterfasern trafen im Pontocerebellum ein und wurden dort nochmals durch sequentielle Divergenz auf mehrere hintereinander angeordnete Purkinjegruppen verteilt.
Damit standen im Pontocerebellum für eine Clustergruppe aus n Cortexclustern wieder n Kletterfasersignale zur Verfügung. Da beide Inputpartner fA und fB höherfrequent waren, war trotz der exponentiellen Dämpfung auch das Überlagerungsergebnis im Divergenzgitter des Nucleus olivaris höherfrequent. Dies war wichtig für die Verwendung dieser tetanischen Signale für die Langzeitdepression und die Langzeitpotenzierung im Pontocerebellum. Doch die erforderte auch Parallelfasersignale.
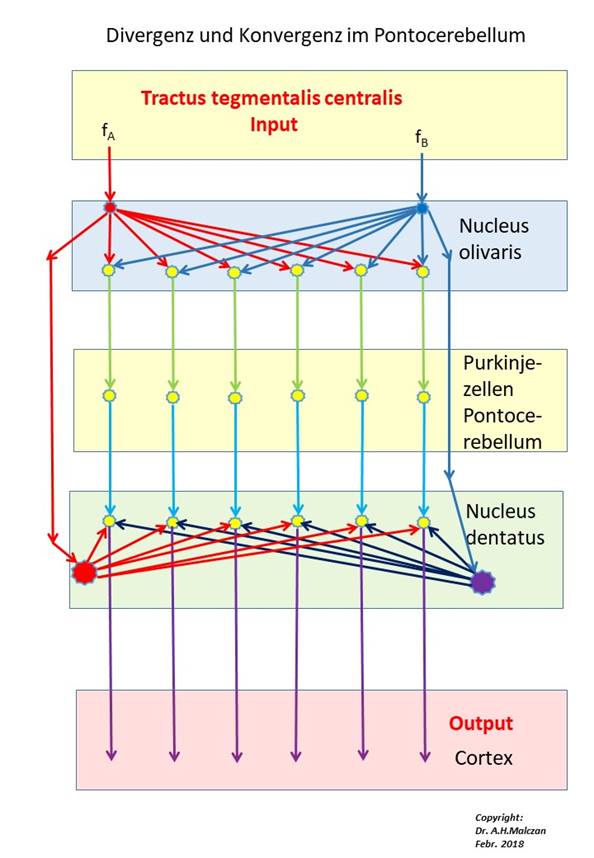
Abbildung 38- Signaldivergenz und Signalkonvergenz im Pontocerebellum
In obiger Abbildung ist nur der Kletterfasereingang zu den Purkinjezellen dargestellt, der Signalzufluss über die Parallelfasern ist (vorläufig) weggelassen. Die Feuerraten fA und fB erreichen nicht nur die zwei Inputneuronen des Nucleus olivaris, sondern über den Tractus tegmentalis centralis - nach Transformation in ein tetanisches und mit Schwingungspausen versehenes Kletterfasersignal - ebenso die zugeordneten Neuronen des Nucleus dentatus.
Neben diesen zwei Feuerraten der beiden Kletterfasersignale trafen im Pontocerebellum auch die Cortexsignale aus den zwei Clustergruppen im Pontocerebellum ein (in obiger Abbildung weggelassen!). Die Cortexsignale verliefen - topologisch wohlsortiert - über die Brückenkerne der Kreuzungsetage zu den Moosfasern des Pontocerebellums. Diese erregten über Kollateralen die Kleinhirnkerne und endeten an den Körnerzellen des Pontocerebellums. Jeder Cerebellumcluster erhielt eine eigene Kletterfaser, seine Purkinjezellen empfingen den Moosfaserinput aus genau einem Cortexcluster.
Clustertheorem des Cerebellums
Die Signalneuronen eines Cortexclusters enden im zugeordneten Cerebellumcluster an den Körnerzellen dieses Clusters. Nachbarschaftsbeziehungen zwischen den Clustern bleiben weitgehend erhalten. Jeder Cerebellumcluster wird von einer Kletterfaser versorgt, die durch Signaldivergenz aus dem Mittelwertsignal der Clustergruppe abgeleitet wird. Dieses Clustergruppensignal des Nucleus subthalamicus wird mit Hilfe des Striosomensystems der Basalganglien in eine getaktete tetanische Schwingung transformiert, die im Nucleus olivaris durch lineare Signaldivergenz auf die Cerebellumcluster aufgeteilt wird. Jede Kletterfaser kontaktiert in ihrem Cerebellumcluster eine Reihe von hintereinander angeordneten Purkinjegruppen, da sie eine sequentielle Divergenz aufweist.
Wenn der Divergenzgrad im Nucleus olivaris beispielsweise gleich vier ist, entstehen so aus jedem Clustergruppensignal vier neue Kletterfasersignale. Jedes von ihnen trifft in einem der vier Cerebellumcluster ein. Zu jedem Cerebellumcluster gehörend nun eine Reihe von hintereinander angeordneten Purkinjegruppen.
Beim Menschen versorgen etwa 430.000 Neuronen des Nucleus subthalamicus 15.000.000 Purkinjezellen. Somit kommen auf ein subthalamisches Neuron etwa 36 Purkinjezellen. Teilen wir diese auf vier Streifen auf, so enthält jeder Streifen 9 Purkinjezellen. Wenn nun drei Purkinjezellen eine Purkinjegruppe bilden, so versorgt eine Kletterfaser 3 Purkinjegruppen. Da jede Purkinjegruppe ein bestimmtes Komplexsignal erlernen kann, würde das Cerebellum etwa 5.000.000 solcher Komplexsignale erlernen können, also 5 Millionen von ihnen. Dies trifft natürlich nicht ganz zu, da ein Teil des Cerebellums als Vestibulo- und Spinocerebellum nur der Signalinversion dient.
Wie viele Cortexcluster könnten in einem Komplexsignal berücksichtigt werden. Eine Parallelfaser beim Menschen erreicht etwa 460 Purkinjezellen, also ungefähr 150 Purkinjegruppen. Wenn je drei Purkinjegruppe zu einem Cortexcluster gehören, bleiben 50 Cortexcluster übrig. Dann könnte eine Purkinjezelle von 50 Cortexclustern über die Parallelfasersignale erreicht werden. Hinzu kommt die Reichweite der Korbzellen. Je nach Literaturstelle kann eine menschliche Korbzelle zwischen 8 und 80 Purkinjezellen erreichen. Damit kann eine Purkinjezelle den Input von 400 bis 4000 Cortexclustern erreichen. Dies entspricht einem cortikalen Rechteck, dessen Kante eine Länge von etwa 20 bis 63 Cortexclustern besitzt.
Wenn wir uns den primären visuellen Cortex anschauen und uns die Orientierungssäulen dazu vorstellen, die in der Cortexrinde zu einer Augendominanzsäule gehören, so könnte man sich diese quadratische Fläche in 400 Teilquadrate unterteilt denken, also in 400 Cortexcluster. In jedem dieser Cluster gibt es dann ein Mittelwertneuron, aus dem das Kletterfasersignal abgeleitet wird. In den zugehörigen 400 Cerebellumclustern könnte eine beliebige Purkinjezelle über die Parallelfasern und die Korbzellen den kompletten Input aus der Augendominanzsäule, also den 400 zugehörigen Cortexclustern empfangen. Jede von Ihnen könnte durch Prägung eine Orientierungsrichtung erlernen. Damit wäre das Pontocerebellum in der Lage, eine Richtungsauflösung von mehr als einem halben Grad zu erreichen, da 180 Grad eine komplette Rotation darstellen. Je länger die Parallelfasern sind und je mehr Purkinjezellen von einer Korbzelle erreicht werden, umso feiner wird das Auflösungsvermögen im Pontocerebellum sein. Daher ist es nicht verwunderlich, dass diese Größen mit fortschreitender Evolution zunehmen.
Wie bisher bereits beschrieben wurde, konnte jede Purkinjegruppe im Cerebellumcluster eintreffende Komplexsignale erlernen, auch schon vor der Signaldivergenz im Nucleus olivaris. Der Algorithmus war der gleiche. Allerdings ist durch die Signalkonvergenz der Mittelwertsignale im Nucleus subthalamicus die Feuerrate der globalen Mittelwertsignale und damit der Kletterfasersignale deutlich erhöht, wodurch LTD und LTP wesentlich effektiver werden.
An dieser Stelle müssen wir die hemmende Projektion des Kleinhirnkerns zum Olivenkern beachten. Jedes glutamaterge Outputneuron des Nucleus dentatus aktivierte (mindestens) ein hemmendes, GABAerges Neuron, welches die laterale Nachbarhemmung realisierte. Ob dieses Neuron gleichzeitig hemmend in den Nucleus olivaris projizierte oder ein eigenes, separates Projektionsneuron mit dem Transmitter GABA aktiviert wurde, um diese Hemmung in der Olive vorzunehmen, ist letztlich egal. Das aktive Outputneuron hemmte im Erkennungsfall über zwischengeschaltete Hemmungsneuronen die Signalquellen im Nucleus olivaris, also die zwei signalliefernden Neuronen, die die Erregungen fA und fB empfingen und divergent auf die Outputzeile des Nucleus olivaris verteilten.
Da dies für jedes Outputneuron des Cerebellumclusters galt, projizierten sämtliche Hemmungsneuronen des Clusters hemmend in genau die zwei Outputneuronen des Nucleus olivaris, die über eine Signaldivergenz sämtliche Kletterfasersignale des Cerebellumclusters bereitstellten.
Diese Hemmung war nicht total, sondern relativ. Die bisherige tetanische Erregung wurde in ihrer Feuerrate so weit abgeschwächt, dass sie niederfrequent wurde und für eine weitere Langzeitdepression oder Langzeitpotenzierung im Pontocerebellum nicht mehr ausreichte. Eine totale Hemmung wäre ungünstig, da dauerhaft unerregte Neuronen möglicherweise dem Untergang geweiht wären. Die verbleibende, niederfrequente Resterregung konnte jedoch durch das Phänomen der Langzeitpotenzierung (LTP) weitere, interessante Resultate bewirken. Es gibt derartige Effekte, die durch niederfrequente Stimulation ausgelöst werden.
Theorem der Hemmung des Nucleus olivaris durch den Nucleus dentatus
In den Kleinhirnkernen bewirkten hemmende Interneuronen eine Kontrastverstärkung des Outputs. Im Nucleus dentatus wurde die laterale Hemmung so perfektioniert, dass das meisterregte Neuron einer Neuronenzeile seine Nachbarneuronen in der gleichen Zeile komplett hemmte. Zusätzlich erregte es ein hemmendes Projektionsneuron, welches zu den beiden Inputneuronen des Nucleus olivaris projizierte, um deren Mittelwerterregung soweit zu hemmen, dass sie keine tetanische, sondern nur noch eine niederfrequente Erregung aufwiesen.
Alle Outputneuronen einer Neuronenzeile waren mit diesem hemmenden Outputneuron verbunden, so dass nun, egal welches von ihnen aktiv war, die tetanische Erregung der zugehörigen zwei Inputneuronen im Nucleus olivaris auf ein niederfrequentes Niveau vermindert wurde. So wurde verhindert, dass ein bestimmtes Komplexsignal im Cluster mehrere Purkinjegruppen prägen konnte, also eine Mehrfachprägung auftrat.
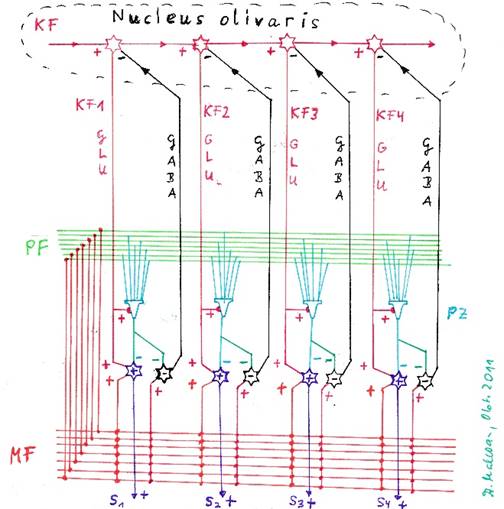
Abbildung 39 - Die Hemmung der Olive durch die Neuronen des Nucleus dentatus
Nun sollten wir die Begriffe Prägungssignal, Eigensignal und Fremdsignal definieren.
Definition: Prägungssignal, Eigensignal, Fremdsignal
Das cortikale Signal, welches in einer Purkinjegruppe über LTD und LTP eine Prägung hervorrief und dazu führte, dass das zugehörige Outputneuron des Nucleus dentatus als einziges mit einem starken Output reagierte, bezeichnen wir als Prägungssignal , den Prozess nennen wir Prägung .
Durch die Prägung wird das Prägungssignal zum Eigensignal der betreffenden Purkinjegruppe.
Jedes von diesem Signal verschiedene Signal bezeichnen wir als Fremdsignal der Purkinjegruppe.
Das Erlernen neuer Prägungssignale ist ein Prozess, bei dem immer mehrere Purkinjegruppen beteiligt sind, die den gleichen Input erhalten. Diese stehen untereinander in Konkurrenz. Diejenige Purkinjegruppe, bei der die Prozesse der LTD und LTP die stärksten synaptischen Veränderungen hervorrufen, geht als Gewinnerin hervor. Die Konkurrenzsituation wird durch mehrere Algorithmen unterstützt.
Einerseits wirken die Kletterfasersignale mit ihrer tetanischen Erregung auf die Golgizellen ein und erregen diese stark. Diese Golgizellen unterbrechen den Signalfluss zu den Körnerzellen, so dass nachfolgend angeordnete Purkinjezellen weniger Input erhalten und ihre Prägung geringer ausfallen muss. So wird innerhalb einer Reihe von Purkinjezellen dafür gesorgt, dass die erste ungeprägte Purkinjegruppe als erste die Prägungsschwelle erreicht und mit ihrem Erkennungssignal das Kletterfasersignal unterbricht. Aber die nachfolgenden Purkinjezellen haben eine unvollständige Prägung erfahren. Diese kann durch weitere Algorithmen im Verlaufe der Zeit wieder rückgängig gemacht werden. Den dazu nötigen Vorgang werden wir hier als Entprägung oder Prägungsumkehr bezeichnen.
Das Phänomen der Prägungsumkehr kann im Cerebellum auftreten, wenn die Purkinjezellen niederfrequent erregt werden und die Parallelfasern gleichzeitig aktiv sind. Als Ergebnis tritt eine Zunahme der synaptischen Kopplung zwischen den aktiven Parallelfasern und der Purkinjezelle auf. Dies setzt jedoch voraus, dass die Purkinjezelle wesentlich länger bzw. häufige aktiviert wird. So wird eine niederfrequente Aktivierung einer Purkinjezelle und die gleichzeitige Aktivierung einer Parallelfaser für mindestens 300 Sekunden benötigt, wie neuere Forschungen [117] von Varda Lev-Ram, Scott T. Wong, Daniel R. Storm und Roger Y. Tsien im Jahre 2002 zeigten.
Wir gehen nun davon aus, dass das cortikale Prägungssignal für eine Gesamtdauer von 300 Sekunden aktiv sein möge, wobei zwischendurch möglicherweise auch Pausen eingeschoben sein könnten.
So wird die unvollständige Prägung der Synapsen zwischen den Parallelfasern und den Purkinjezellen rückgängig gemacht, indem der durch LTD erniedrigte Kopplungsgrad durch LTP wieder erhöht wird.
Für die Synapsen zwischen den Korbzellen und den Purkinjezellen bzw. den Sternzellen und den Purkinjezellen, deren Kopplungsgrad durch unvollständige LTP vergrößert worden war, gibt es einen analogen Effekt der Langzeitdepression durch niederfrequente Kletterfasereinwirkung.
Die unvollständige Hemmung der Kletterfasersignale beim Erkennen des Eigensignals einer Purkinjegruppe dient der Rückgängigmachung der unvollständigen Prägung bei den Nachbargruppen im Cerebellumcluster.
Theorem der erkennenden Purkinjezelle
Erkennt eine Purkinjegruppe im Pontocerebellum ihr cortikales Prägungssignal, so hemmt ihr zugeordnetes Dentatusneuron über ein zwischengeschaltetes Hemmungsneuron sofort das mit zeitlicher Verzögerung eintreffende Kletterfasersignal, so dass es niederfrequent wird und für eine erneute Prägung im Pontocerebellum nicht mehr zur Verfügung steht. Das nunmehr niederfrequente Kletterfasersignal ruft in den benachbarten Purkinjezellen, die unvollständig geprägt wurden, eine (langsame) Entprägung des aktuellen Prägungssignals hervor, indem zwischen den aktiven Parallelfasern und den Purkinjezellen der Kopplungsgrad durch LTP bis zum Wert 1 erhöht wird. Für die Synapsen der Korb- und Sternzellen mit den entsprechenden Purkinjezellen wird der Kopplungsgrad auf den Ausgangswert vor der teilweisen Prägung erniedrigt. So wird letztlich nur eine Purkinjegruppe auf dieses Prägungssignal reagieren.
Was aber passiert, wenn nicht das Prägungssignal, sondern ein davon verschiedenes auf die geprägte Purkinjezelle einwirkt?
Wesentlich für die Phänomene LTD und LTP ist der zeitliche Vorsprung, mit dem die Signalklasse 5 vor der Signalklasse 6 im Pontocerebellum eintrifft. Während die cortikalen Signale der Klasse 5 über die Kreuzungsetage und die Moosfasern bei den Körnerzellen eintreffen und die Parallelfasern aktivieren, müssen die Mittelwertsignale der Klasse 6 den wesentlich längeren Weg über die Substantia nigra pars compacta, das Striatum, den Nucleus ruber und den Nucleus olivaris nehmen. Daher treffen sie deutlich zeitverzögert ein.
Trifft also ein vom Prägungssignal verschiedenes Signal ein, dann wird diese Purkinjezelle von den eintreffenden Cortexsignalen stark erregt, denn die synaptische Kopplung zwischen den Parallelfasern und der Purkinjezelle hat den Wert 1. Diese starke Erregung wird aufgebaut, bevor das Kletterfasersignal über den Tractus tegmentalis centralis eintreffen kann. Die starke Erregung der Purkinjezelle hemmt das zugehörige Outputneuron im Nucleus dentatus völlig. Es gibt keinen Output, solange das Kletterfasersignal seine Austastpause hat.
Trifft nun das Kletterfasersignal mit zeitlicher Verspätung ein, so ist es prinzipiell wirkungslos. Die von der Purkinjezelle verursachte starke Hemmung bewirkt einen Kurzschluss zwischen dem Außenraum und dem Innenraum der Zelle im Nucleus dentatus, die Hemmung setzt sich gegen die Erregung durch. Daher liefert das Outputneuron keinen Output. Dies gilt für alle Purkinjezellen der betreffenden Purkinjegruppe.
Theorem der Reaktion auf ein Fremdsignal
Eine geprägte Purkinjegruppe reagiert auf das Eintreffen eines Fremdsignals mit einer starken Hemmung des zugehörigen Outputneurons im Nucleus dentatus. Dies ruft einen Kurzschluss in dem Outputneuron hervor, so dass die verspätet eintreffende Kletterfasererregung wirkungslos bleibt und kein Output auftritt.
Besteht jedoch ein Komplexsignal teilweise aus dem Eigensignal einer Purkinjezelle und besitzt weiterhin auch Anteile eines Fremdsignals, so wird die Purkinjezelle vom Eigensignalanteil weniger erregt als vor der Prägung, die Hemmung des Outputneurons wird vermindert und es gibt einen Output. Dieser fällt jedoch geringer aus, als wenn nur das Eigensignal vorhanden gewesen wäre. Dies ist ein typisches Verhalten von neuronalen Netzen.
- ← 4.2 Die zweite Erweiterungsphase des urtümlichen Spinocerebellums
- 4.4 Die Aufspaltung des Neuralrohrs während der Cerebellumentwicklung →
Monografie von Dr. rer. nat. Andreas Heinrich Malczan