Gehirntheorie der Wirbeltiere
ISBN
978-3-00-064888-5
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
Die Entstehung von Divergenzgittern im Cortex
Motorische Divergenzgitter zur Körperüberwachung und Steuerung
Im Fachbuch -Neurowissenschaften- [1] von Eric R. Kandel, James H. Schwartz und Thomas M. Jessell, Spektrum Akademischer Verlag 1995, werden im Kapitel 29 die Forschungsergebnisse zur Willkürmotorik dargestellt. Es wird gezeigt, dass Neuronen im primären motorischen Cortex die Kraft und die Richtung von Willkürbewegungen kodieren.
In einer Versuchsanordnung bewegten Affen einen Stab mit dem Arm zu einem von mehreren Zielpunkten, die in unterschiedlichen Richtungen um eine zentrale Startposition angeordnet waren.
Während der Bewegungen wurden die Feuerraten von sehr vielen cortikalen Neuronen in dem Gebiet gemessen, in dem die Signale der Rezeptoren der zugehörigen Muskeln und Gelenke eintrafen.
Es zeigte sich, dass die Bewegungsrichtung nicht von einzelnen Neuronen, sondern von Neuronenpopulationen codiert wird. Diese Neuronenpopulationen sind etwa kreisförmig um ein Zentrum angeordnet. Je nach Bewegungsrichtung ist jedoch immer nur eine Neuronenpopulation aktiv. Deren Lage zum Zentrum codiert die Bewegungsrichtung derart, dass der Populationsvektor weitgehend mit der Bewegungsrichtung übereinstimmt.
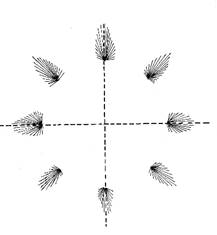
Abbildung 57- Kodierung der Bewegungsrichtung durch Neuronenpopulationen
Quelle: Abb. 57 nach [1], Seite 545, Abbildung 29.3
Und hier erinnern wir uns an die Erregungsfunktion in ebenen Divergenzgittern. Ein Gelenk wie das Schultergelenk ermöglicht es, den Arm beliebig rotieren zu lassen. Vereinfachend ist eine Muskelgruppe für das Heben des Arms nach vorn zuständig. Eine weitere für das Absenken. Beide wirken gegeneinander und liefern das erste Paar komplementärer Signale. In diesen verschlüsselt die Urgröße u den Winkel der Armhebung nach vorn.
Eine weitere Muskelgruppe ist dafür zuständig, den Arm seitlich zu heben, die zugehörigen Gegenspieler senken den Arm seitlich wieder ab. Beide wirken wieder gegeneinander und liefern das zweite Paar komplementärer Signale. In diesen verschlüsselt die Urgröße v den Winkel der seitlichen Armhebung. Das Zusammenwirken der vier Muskelgruppen ermöglicht die Armbewegung in beliebige (anatomisch zulässige) Richtungen.
Es ist nicht schwierig, den Urgrößen
u und v eine Darstellung in Polarkoordinaten zuzuordnen. Mit r² = u² + v² und
![]() sowie
sowie
![]() erhält
man den Radiusvektor r und den Rotationswinkel φ. Eine kreisende Armbewegung
würde in Polarkoordinaten einen kreisenden Radiusvektor r ergeben. Je größer der
durch die kreisende Armbewegung beschriebene Kreis wäre, umso größer wäre auch
der Radiusvektor r in Polarkoordinaten.
erhält
man den Radiusvektor r und den Rotationswinkel φ. Eine kreisende Armbewegung
würde in Polarkoordinaten einen kreisenden Radiusvektor r ergeben. Je größer der
durch die kreisende Armbewegung beschriebene Kreis wäre, umso größer wäre auch
der Radiusvektor r in Polarkoordinaten.
![]() (4.1.1)
(4.1.1)
![]() (4.1.2)
(4.1.2)
Die zugehörigen Muskelspannungsrezeptoren der beteiligten vier Muskeln (bzw. Muskelgruppen) geben die Erregungen an die Inputneuronen des ebenen Divergenzgitters im motorischen Cortex weiter, die Erregungen überlagern sich und besitzen ein Erregungsmaximum, welches wie der Radiusvektor r rotiert, wenn man die Arme kreisen lässt. Die Länge des Radiusvektors wird von der Größe der Armkreise bestimmt.
Das Erregungsmaximum im Punkt P(xo, yo) ist über die folgenden Gleichungen (3.14) und (3.15) an die Urgrößen u und v gekoppelt:
![]() (3.14)
(3.14)
![]() (3.15)
(3.15)
Die Neuronen sind jedoch nicht nur im Erregungsmaximum erregt, sondern in der näheren Umgebung ebenso, auch wenn die laterale Hemmung dafür sorgt, dass weiter entfernte und damit erregungsschwache Neuronen stärker gehemmt werden und daher fast gar nicht feuern.
Rotiert der Arm, so rotiert auch das Erregungsmaximum und die darum befindliche aktive Neuronenpopulation um den Koordinatenursprung. Wobei nicht die Neuronen rotieren (sie sind ortsfest!), sondern der Ort mit der stärksten neuronalen Aktivität. Und in diesem Ort befindet sich halt nicht ein einzelnes Neuron, sondern eine ganze Population von Neuronen. Dieses real beobachtete Phänomen der Verschlüsselung einer Bewegungsrichtung durch eine maximumcodierte Neuronenpopulation beweist, dass es im Cortex in diesen Gebieten real existierende Divergenzgitter gibt, die sich so verhalten, wie die hergeleitete Theorie es voraussagt.
Das Besondere an dieser Theorie ist, dass sie keinerlei Lernvorgänge voraussetzt. Die Erklärung der beschriebenen Phänomene unter Verwendung von lernenden neuronalen Netzen mit plastischen Synapsen kann zwar ähnliche Resultate liefern. Aber beim Vergleich zweier verschiedener Erklärungstheorien wird häufig derjenigen Theorie der Vorzug gegeben, die die einfachste Erklärung liefert. Das Wirken gewöhnlicher Naturgesetze bei der Ausbreitung neuronaler Erregungen auf marklosen Fasern reicht völlig aus, und die Phänomene in den motorischen Cortexregionen zu erklären. Lernvorgänge in neuronalen Netzen sind dazu überflüssig.
Und bedenken sollten wir auch, dass solche Divergenzgitter bereits bei einfachsten Lebewesen wie etwa Insekten vorhanden sind. Entstanden durch Signaldivergenz - also der Weiterleitung von neuronalen Signalen auf mehreren Axonen. Der Output solcher Divergenzgitter erlaubt die einfache motorische Steuerung - wie sie beim Insektenflug beobachtet wird, ohne dass höhere Gehirnleistungen erforderlich wären. Zumal einige extrem kurzlebige Insekten gar keine Zeit hätten, neuronale Netze mit riesigen Datenmengen zu füttern, damit sie Bewegungsmuster erlernen.
Ursache für die Extremwertcodierung ist die nichtlineare Signaldämpfung auf marklosen Fasern. Wir hatten in dieser Monografie spezielle Annahmen zur Signaldämpfung gemacht. Es ist jedoch durchaus möglich, auch andere Annahmen zu machen, andere Funktionsgleichungen zu wählen. Die Ergebnisse werden analog sein. Die Lage von Extremwerten wird (fast) immer eine stetige Funktion der Funktionsparameter sein, und Letztere sind die Ortskoordinaten der beteiligten Neuronen. Man könnte daher untersuchen, welche Funktionenklassen zu analogen Resultaten führen.
Theorem der Maximumcodierung von Bewegungsrichtungen in ebenen Divergenzgittern
In ebenen Divergenzgittern im motorischen Cortex wird die Bewegungsrichtung durch einen maximumcodierten Populationsvektor verschlüsselt. Ursache ist die Signaldämpfung bei der Ausbreitung von neuronalen Erregungen.
Hierbei muss gesagt werden, dass die Aktivität der Neuronen im Populationsvektor zunächst das Resultat der eintreffenden Rezeptorsignale darstellt. Hier darf Ursache und Wirkung nicht verwechselt werden. Ursache sind die Rezeptorsignale und die damit verbundenen Muskelbewegungen, die Wirkung ist die Ausrichtung des Populationsvektors entsprechend der Bewegungsrichtung.
Anders wäre es, wenn andere Gebiete der Großhirnrinde ihre Signale in das ebene Divergenzgitter senden würden. Dann käme es ebenfalls zur Ausbildung eines Populationsvektors. In diesem Falle jedoch würde der Output des Divergenzgitters - nachdem er ein zugehöriges Konvergenzgitter durchlaufen hätte - zur Muskelsteuerung verwendet. So könnten Gedanken die Bewegungsrichtung steuern.
Die Entwicklung der Signaldivergenz in den primären Cortexgebieten, die im Urhirn der Bilateria durch die oberste Etage des Strickleitersystems repräsentiert wurden (die sich bekanntlich in verschiedene Modalitäten aufgespalten hatten), hatte weitreichende Folgen.
Die aufsteigenden Axone aus dem Neuralrohr bzw. Rückenmark erreichten über den Thalamus der zugehörigen Modalität (die zweite Etage des urtümlichen Strickleitersystems) den Cortex und dockten dort an den Neuronen der Klasse 4 an. Diese waren nun lokale Interneuronen. Sie übertrugen ihre Erregung an die Neuronen der Klasse 3, die im Strickleitersystem waagerecht zum motorischen Leiterholm projizierten. Als Folge der zunehmenden Signaldivergenz nahm die Anzahl der Neuronen der Klasse 3 enorm zu. Die zugehörige Fläche dehnte sich zunehmend aus und bildete eine Art Kugelsegment. Bei Raummangel trat Faltenbildung auf, wodurch sich die verfügbare Fläche deutlich vergrößern ließ. Die zugehörigen Axone wurden so zahlreich, dass sie nicht mehr waagerecht zum motorischen Leiterholm verlaufen konnten, zumal auf der motorischen Seite eine ebenso starke Zunahme der Empfangsneuronen der Klasse 5 stattfinden musste. So wurde die Außenfläche des Cortex kugelartig, und in ihrem Inneren verliefen die Axone der Neuronen der Klasse 3 vom sensorischen zum motorischen Teil. Dort trat nun die notwendige Signalkonvergenz auf, spezielle Neuronen der Klasse 5 (Betzsche Pyramidenzellen) empfingen den Output des zugehörigen Divergenzgitters und hoben die Signaldivergenz wieder auf. So erhielt das Neuralrohr bzw. das Rückenmark die durch Signalkonvergenz deutlich verringerte Anzahl der motorischen Outputsignale.
Die cortikalen Divergenzgitter erlaubten eine viel bessere Auflösung von Signalen. Gelenkwinkel konnten dargestellt werden und in den Assoziationsgebieten viel feiner mit anderen, ebenfalls genauer aufgelösten Signalen kombiniert werden. Das ursprüngliche Ja-Nein-Prinzip, entweder feuern oder schweigen, wurde abgelöst durch eine viel präzisere Bewertung von Signalen über maximumcodierte (bzw. extremwertcodierte) Signalpopulationen. Dies erlaubte eine viel feiner gesteuerte Motorik, war jedoch auch eine Voraussetzung für die Entwicklung höherer intellektueller Leistungen. Die durch Signaldivergenz entstandenen, extremwertcodierten Signale erreichten über Neuronen der Klasse 5 auch das Basalgangliensystem und das Pontocerebellum zur weiteren Auswertung.
- ← 5.4 Theorie der ebenen Konvergenzgitter
- 5.6 Die Richtungsselektivität der visuellen Divergenzgitter →
Monografie von Dr. rer. nat. Andreas Heinrich Malczan