Vertebrate brain theory
ISBN
978-3-00-064888-5
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
3.22 The analogue primal brain of the early choral data
The analogue primal brain of the early choral dataThe primeval brain of the early choral data described above realized its entire signal processing in analog form. The rate of fire was always the decisive analog quantity. On the one hand, this was true for the transformation of sensory signals into motor control signals, as was the case in the torus semicircularis or the tectum opticum.
Similarly, the analogous mode of operation was realized in the initial contralateral inhibition and later in the inverse excitation of the motor opponents. The vestibulo- and the (early) spinocerebellum worked analogously via the rate of fire.
The analog fire rate was also the decisive factor in motion detection in the basal ganglia system and the amygdala.
Even the ancient Pontocerebellum based his ability to learn exclusively on analog signals.
We can therefore rightly say that the early primeval brain of choral data worked in an analogous way. We will also use the term primary vertebrate brain for the analogous primal brain.
Theorem of the analogous primal brain
The early primeval brain of choral data worked analogously.
However, this statement only makes sense if there are other ways of working of the chordal brain that are not analogous. In this case, the working hypothesis must be put forward - even against the bitter resistance of many experts - that two further working methods developed in the vertebrate brain in the course of further evolution. This led to the formation of new substructures that can be summarized organizationally. Finally, the vertebrate brain is divided into three parts: a primary subbrain, a secondary subbrain and a tertiary subbrain. The primary subbrain uses analog signals, the secondary subbrain uses digital signalsand the tertiary subbrain uses complex signals. And since the author proves this assertion step by step, the critics must take the trouble to prove serious errors in this presented evidence. It is unacceptable to simply ignore this theory in order not to have to refute it! In this respect, the experts of the European Union's Human Brain Project have a moral obligation to take note of and examine this vertebrate brain theory.
The vertebrate brain is - functionally but not spatially - divided into three parts: a primary subbrain, a secondary subbrain and a tertiary subbrain. The primary subbrain uses analog signals, the secondary subbrain already digital and the tertiary subbrain complex signals. The primary subbrain is spatially diffusely distributed in the secondary subbrain and only occasionally forms independent spatial substructures such as the striosomes in the matrix, the magnocellular part in the nucleus ruber or the Betzs pyramidal cells in the motor cortex. It is virtually hidden and is only noticeable in very early forms of vertebrates. The tertiary subbrain is fully developed in Homo sapiens, but less developed in other species. Nevertheless, its beginnings go back to early primeval times.
Each of these three structures has an independent signal form. Each of the three structures exchanges its signals with the other two subsystems. The previous signal form (analog, digital, complex) must first be transformed into the new signal form of the receiver system. These signal transformations are performed by transformation systems. Among them are the cerebellum, but also parts of the sensory cortex, the basal ganglia and the limbic system.
Similar transformation structures must be passed through when the signals of one subsystem are passed on in descending order to the other subsystems. Therefore there are ascending and descending projecting transformation systems. Among the descending projecting systems are the nucleus ruber, but also the motor cortex or the dorsal ventricular crest (DVR) in reptiles or birds.
Each of the three structures has its own short-term memory in the form of a runtime system in which a small time delay is imposed on the signals as they are transported on non-markless axons. This runtime system enables the signal comparison between undelayed and delayed signals in the form of a difference mapping, which is mainly used for motion detection.
Furthermore, each of the three structures has a separate rotational memory in which the signals rotate permanently, thus keeping alive the memory of the past, which was already long ago. Periodically every 24 hours - or earlier if necessary - the rotating signals can be deleted. The nucleus suprachiasmaticus and other time-controlled clocks are integrated here.
The tertiary system has a complex signal memory that can store important signals and recognize them later. Here there is a neuronal storage command as well as a recognition signal as output, if a signal is recognized.
The output of the tertiary system has again the analog data format and is recursively fed back into the primary system. This is no longer dealt with in this monograph; the interested reader should refer to earlier monographs by the author. The signal rotation in the brain system described in an earlier monograph is not dealt with either, as this is primarily about the vertebrate brain, not so much about the brain of Homo sapiens. We have to limit ourselves a little here in terms of the subject matter.
Initially, a digitally working subbrain developed above the analogue primordial brain, which was later to become a complex subbrain. Modern vertebrates therefore have an analog, a digital and a complex subbrain. The way to the digital subbrain led through the second expansion phase of the spinocerebellum, a parallel development took place in the substructures of the amygdala. This is described below.
However, the complex subbrain does not work with complex numbers, as one might assume, but works with complex signals. This term has been used in neurology for a long time and retains its previous meaning in this monograph, only that it becomes mathematically describable.
It takes some getting used to the digital format in the vertebrate brain. There are no numbers. You have to imagine the numerical values as represented on a number ray with the natural numbers, but the set of numbers is finite. An output neuron is assigned to each numerical value. The actual numerical value is represented by the activity of a group of neurons, which we call extreme value coding. This is possible in two ways. With maximum value coding, the numerical value is encoded as the maximum in the active neuron group, the other neurons are inactive. With minimum coding, however, all neurons are active with the exception of a neuron group that contains an absolute minimum excitation.
This type of encoding is unusual, but offers a great advantage: If one neuron fails, a neighboring neuron takes over the extreme value encoding. In this way, neuronal circuits achieve an unmatched reliability.
An overview of the structural division of the vertebrate brain into the primary, secondary and tertiary subbrain is given in the following figure.
The input and output cores are not drawn in, as otherwise the display would have become unclear. At the beginning of each arrow you have to imagine an output core, whose neurons receive the input and guide it with their axons to the target system. There the neurons of the input core of this substructure receive the signals. In principle, 13 subsystems have 13 input cores and 13 output cores. However, the signals of different modalities usually use their own subsystems. For example, the different lobi of the brain process different modalities, as do the olfactory and limbic systems. As a result, the number of neuron nuclei required to function as input or output nuclei is significantly larger. For the sake of clarity, they have therefore been omitted in the following overview.
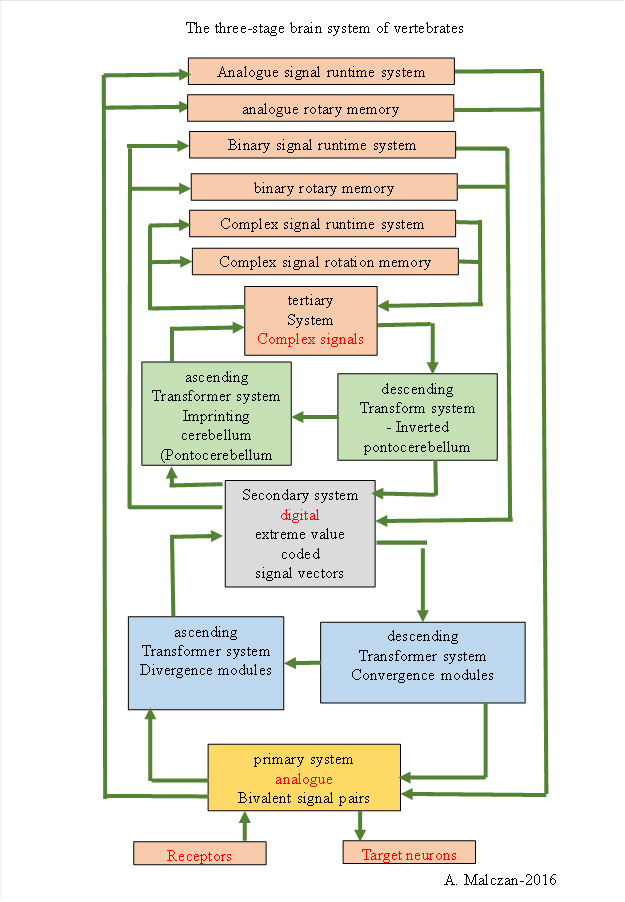
Figure 22 - Schematic diagram of the three-part vertebrate brain according to A. H. Malczan
All essential components of the analogue primeval brain have been described in the previous chapters. Now we turn to the digitally operating subbrain.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan