Vertebrate brain theory
ISBN
978-3-00-064888-5
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
3.3 The specialization of the head segments and their merging
We divide the body of the segmented Bilateria, whose line led to the vertebrates, into a trunk area and a head area. The head region includes a certain number of cranial segments. In segmented animals living today, the head region consists of approximately four to seven segments, depending on the species. The special feature is that the segments lying in front of the actual mouth area are partly fused together.
Due to the ladder-like structure of the nervous system and the ascending projections in the sensory ladder spar, in each segment not only the excitation was available, which was transmitted by the receptors of this segment to the sensory centre, but also the signals of all receptors, which were obtained in the deeper, tail-side segments. Between all these signals a lateral inhibition could occur due to inhibiting interneurons. As a result, not only the left and right halves of the body were in neuronal competition with each other, but also the individual segments. This led to a special development in the interdependent receptors.
Theorem of neuronal competition between segments
Since in each segment the signals of all subordinate segments were available and the stronger signals inhibited the weaker ones via the lateral inhibition in the respective neuronal nucleus, and since there was contralateral inhibition of the corresponding neuronal nuclei at each segment level, all segments of the body were in neuronal competition with each other.
The receptors of the different segments can be divided into two groups according to their modality. Dependent receptors deliver approximately the same signal in each segment. Related to a longer period of time, the signals of dependent receptors (statistically seen) differ only very little. For example, scent receptors can detect a scent evenly distributed in water with approximately the same intensity in each segment of the body. We call such receptors dependent receptors (in analogy to linearly dependent vectors). If, on the other hand, the receptors of one type of receptor in each segment provide independent signal values, because the muscle tension of the muscles in different segments (usually) differs, we call these receptors independent.
In the course of evolution a specialization of the dependent receptors of the different segments occurred. If a receptor type in a segment had produced an above-average number of receptors, this segment could specialize in the type of stimulus (modality) detected by this receptor type. This is because the signals of this segment were significantly stronger compared to the signals from the other segments of the same type of receptor. They suppressed the similar signals from the other segments that reached this segment via class 4 neurons and deactivated them permanently via the lateral neighbor inhibition. This leads to the regression of this type of receptor in the remaining segments. Thus, this segment specialized on a certain type of receptor.
It was also possible, however, that several types of receptors in one segment dominated simultaneously compared to the same receptors in other segments.
For the independent receptors, such a development could not take place because the lateral neighbor inhibition was only partial and there were sufficient phases for each receptor in each segment in which no lateral inhibition occurred due to missing neighbor signals.
Among the dependent receptors we count the olfactory receptors, the receptors of the sense of balance, the visual receptors, the taste receptors. These types of receptors have concentrated on individual segments in the course of evolution. This occurred preferentially in the segments that we count as the head area. Lateral inhibition was most effective there, and only there were the signals of the entire body available.
Among the independent receptors we count those of the lateral line sense, the muscle tension receptors (initially free nerve endings, later muscle spindles), the receptors of the tendon organs as well as those of the sense of touch and pain. The receptors of the animal system are not classified here.
This resulted in a dichotomy of the receptors: In the head area, the dependent receptors gained dominance via lateral inhibition and led to the specialisation of the head segments on different receptor types.
Theorem of sensory specialization of the head segments
The neuronal competition between the receptor signals of the different segments led to the fact that dependent receptors were ultimately only represented in a single head segment. Therefore, the different head segments specialized in the perception of stimuli of different modalities. Thus, each modality in the head segment was processed in only one segment, even though one head segment could well be responsible for several modalities.
Initially, all segments had the same equipment of organs, only the digestive system, blood circulation and nervous system were shared.
However, since the organs were controlled by nerve signals via the mean value centres, the neuronal competition between the segments also led to a reduction here, so that in the course of evolution, not every organ was present in every segment, but only once per half of the body. Here, it could possibly overcome the segment boundaries due to its increasing size. If an organ in a certain segment was no longer sufficiently supplied with input signals due to the neuronal signal competition between the segments, this led to a reduction in its size and ultimately to regression. Therefore, the initially completely identical segment structure was abandoned in the course of the longer evolutionary period and the segments differentiated and divided the tasks among themselves. An example for the regression of an organ is nervous atrophy. If a nerve that supplies a muscle is damaged, this muscle, which is no longer supplied, atrophies. Analogously, we can imagine the regression of organs that were initially present in every segment of the body. Due to the lateral inhibition of the supplying nerves in the neuronal centers, a competition among these organs arose, as a result of which individual segments were less strongly innervated, which further accelerated the lateral inhibition. Thus, organs gradually regressed until this type of organ was left in only one segment and was unrivalled there.
When this gradual change in DNA during evolution was recorded and passed on to the descendants, there were no longer multiple organs (with the exception of those that appeared in one segment on each side of the body due to bilateralism. A prerequisite for such a development was of course the existence of inhibitory interneurons that realized the lateral neighbor inhibition.
Theorem of successive organ regression in multiple organs
Lateral neuronal inhibition caused competition between the organs present in (almost) every segment in the initial phase of segmented animals. This led to a reduction in the number of these organs, so that they ultimately remained in only one segment.
In later evolutionary times, the various sensory and motor centers of the head area merged, a development we call the fusion of the head segments.
It can be assumed that the lateral inhibition in the neuronal centers was not developed to the same extent in all segmented bilateria. There were certainly animal species among them in which the signal range on the afferent or efferent projection neurons of classes 4 and 5 was small, for example because they did not have myelin. In such cases, the signals of the more distant segments could not reach the neuronal centers at all, causing lateral inhibition there. It was also possible that various types of segmented bilateria developed no or too few inhibitory interneurons in their neuronal centers, so that sufficient lateral inhibition did not occur there either. These animal species were not able to gradually regress their organs located in each segment at all, since there was no neuronal competition among them. In their case, these organs remained intact in all segments (possibly with the exception of the head and tail segment). In fact, this original structure of almost identical segments has been preserved in a number of animals to this day (e.g. in worms). In insects, on the other hand, the described regression of multiple organs and a specialization of the segments occurred, which we can also still observe today. Therefore, this phenomenon of successive regression of multiple organs does not only occur in segmented bilateria, whose lines lead to vertebrates, but can be observed in many animal species.
We assume that (with reference to the later vertebrates), in the course of the specialization of the different head segments in different modalities, the class 3 neurons in each head segment ultimately only transmitted signals of that modality to the motor side of the rope ladder system in which this head segment had specialized. This was the more economical variant. It was a prerequisite for the thalamic structures also being able to specialize in selected modalities later on as the head segments merged.
In the course of specialization, the sensory centers of the different head segments therefore became responsible for different modalities. A motor specialization of the different head segments for different functions took place in parallel. The sensory and motor centres of the head segments later became the nerve centres of the brain. All vertebrates have twelve cranial nerves. At the early stage of development described here, however, not all the specifications of the future vertebrate body had been developed, so there was no head skeleton, no jaws, no masticatory muscles, etc. Therefore, many of the later cranial nerves were missing at this early stage of development. However, the presence of 12 cranial nerves in vertebrates would suggest that there may have been 12 primal segments that served as head segments. However, it is more likely that some head segments produced multiple modalities, so that their total number would be less than 12.
In vertebrates, the sensory and motor centres of the trunk gave rise to the spinal ganglia.
Theorem of specialization of the first segment on olfactory
The first segment - the head segment - takes the lead in olfactory receptors. The signals of the olfactory receptors located directly in front of the mouth area in the first segment were strongest, they suppress the olfactory signals in the other segments via the lateral inhibition, so that the olfactory receptors there recede and ultimately disappear.
The mean center of the first segment developed into a neuronal nucleus called a septum. It mainly receives olfactory signals from the first segment and ascending signals from the second segment. This mean nucleus was initially used for motor activation and therefore used the transmitter acetylcholine, which causes contraction in the muscle plate. The descending, motor signals were imprinted with the additional activation of the (initially) olfactory mean centre, scent signals therefore enhanced motor activities.
The strong development of various olfactory receptors in the head segment led to the reduction of other sensory receptors, so that the head segment specialized in olfactory science.
The motor centre of the head segment developed into a neuron nucleus called the amygdala.
From the sensory center of the head segment, the hippocampusdeveloped over a longer period of time, which produced the olfactory rotational memory. This will be described later.
With regard to the physical shape, it can be assumed that the olfactory receptors were initially located on small stems and thus extended a little further into the space to be analysed. In vertebrates, for example, the olfactory receptor surfaces are still positioned somewhat eccentrically and protruding forward.
We recall the hypothesis that the first segment specialized in olfactory science. The sensory centre of the first segment directly received the signals of olfactory receptors. The commissure neurons transmitted the signals directly to the downward projecting connective neurons.
We assume that the later cortex was formed from the first floor, while the thalamus structures were formed on the second floor. This is consistent with the fact that the olfactory system projects directly into the cortex and its signals do not take the path via the thalamus (or one of its nuclei).
Theorem of specialization of the second segment on visual signals
The second segment specializes in visual signals. On each side of the body, a compound eye was created, the surface of which was convexly curved, resulting in an upright and laterally correct image of the visual objects on the associated receptor surface, the later retina.
It is possible that initially light-sensitive cells were present at the tip of small stems on the left and right, similar to the stem eyes of snails. Muscle groups could bend these stem eyes in different directions. They (possibly) gave rise to the eye muscles of vertebrates. The number of light-sensitive cells increased in the course of evolution, and a receptor surface was created from which the later retina developed. The stalk on which the eyes sat led the axons of the light receptors to the second segment, where they docked with the neurons of the sensory centre. This basic structure may explain why the eyes of modern vertebrates are located quasi externally and quite far from the actual brain.
Theorem of specialization of the third segment on vestibular signals
The third segment specializes in vestibular signals.
The vestibular sense was based on hair cells that generated action potentials when turned by external forces. Even in the simplest creatures, these hair cells were found in a fluid-filled body cavity, the statocyste, which they lined. At that time, all animals lived in water. The cavity was connected to the surrounding water through a small opening. Through it, a small grain of sand could penetrate into this statocyste. Therefore, the statocyste was located very close to the body surface, partly protruding a little bit. Possibly, the entrance channel was narrowed after the penetration of a grain of sand, so that it could not fall out.
The sand grain was freely movable in this cavity and rolled to its lowest point under the influence of gravity. There its weight caused the hair cell underneath to bend, which then generated action potentials. Depending on the body's inclination, a different hair cell responded in each case. Their signals reached the sensory centre and there led to the excitation of associated neurons. In the end, they generated corrective movements of the living being.
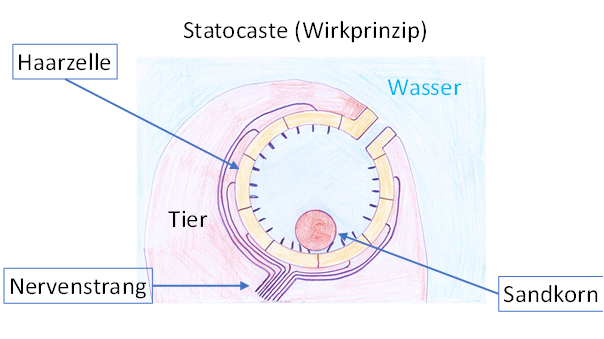
Figure 3 - Statocyste (active principle)
Interested readers can find out more about the sense of balance in the excellent reference book "Tierphysiologie" by Heldmaier, Neuweiler and Rössler [46]. There, in chapter 14.4, the stato system of aquatic invertebrates and vertebrates are presented as examples and their neuronal networks are analysed. The vestibularly triggered corrective movements are illustrated using the example of a marine snail. There it says on page 605:
"Clione limacina is a tiny winged snail (pteropoda) that belongs to the marine plankton. It stands vertically in the water with its head up and compensates any deviation from this vertical position by counteracting tail (i.e. the foot of the snail) movements. Two statocysts, whose wall is lined with 10 hair cells each, use a statolith to measure deviations from the vertical body position. This bilateral position information is switched to 20 specific neurons in the cerebral ganglion. Six of these neurons innervate tail motoneurons in the pedal ganglion."
It is described how this small neuron network including statocysts was prepared from the animal and how its function was demonstrated experimentally.
The following figure - based on the illustration in the book mentioned above - illustrates the correction movements of this wing auger in case of a deviation from the vertical. If an external force acts on the marine snail and rotates it by the angle ß, the vestibular signals cause a curvature of the tail. Due to the water resistance, the snail then regains a vertical orientation.
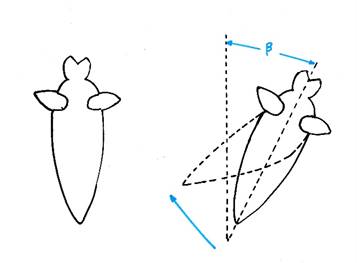
Figure 4 - Vestibular-triggered correction movements
We refer to this vestibular sense in this monograph as paleovestibular sense. It is the oldest way of analysing the effect of the earth's gravitational field on living beings by means of receptors.
Most readers will agree without hesitation that the vestibular sense serves the correct alignment of the body in the earth's gravitational field. Certainly this view may seem correct, especially when it concerns man. For the upright walk is considered a great step forward in evolution. Without a functioning vestibular sense, standing upright, walking or running is almost impossible.
Nevertheless, the very early vestibular sense of the simplest living things and also of the later aquatic vertebrates has been the control system for aquatic locomotion.
Let's just think of a fish with fins that serve to correct the situation. If the body tilted to the left, the right fin corrected the deviation. By the vestibular triggered fin movement the body received an angular momentum and rotated around the longitudinal axis. And this rotation did not simply stop when the body reached the desired standard position in the water. Following the law of inertia, it continued to rotate, because the friction in the water absorbed only a small part of the angular momentum energy. Thus the corrective movement shot past its target. This was immediately detected by the vestibular system and a correction signal was sent out. This activated the contralateral fin to a correction movement. But this also shot over the target and caused a corrective movement in the opposite direction. Thus the body oscillated back and forth between these two deviation positions.
But because the fins were attached to the body (and for other reasons, which we will omit here), every fin movement always produced a propulsion component in addition to a rotational component. Thus the oscillating corrective movements led to propulsion in the water. And this at a time when there were no neural pacemakers. The latter required components that were not available in the elementary bilateria system.
The vestibular sense was one of the first senses besides the olfactory sense and enabled the mobility of the early, non-sessile species. Since there was no brain in the beginning, movements had to be caused by some principle. The vestibular system was able to perform this function very early on. And those species which, in addition to a trunk, also had paired fins, which were controlled by the vestibular signals to correct their position, won the competition for the highest possible propulsion speed. The paired extremities of the vertebrates served from the beginning of the locomotion, and the vestibular sense generated the propulsion signals.
Bilaterally constructed organisms have two statocysts, one on the left and one on the right side of the body. We assume that in the development of segmented bilateria the statocysts were no longer needed in each body segment, since their signals were (almost) identical to those of the third segment. A regression would have been possible, but also a modification. We assume that the hair cells in the statocysts of the subsequent segments changed in such a way that the opening through which a small grain of sand could penetrate became larger over time. Any grains of sand that were present fell out and the enlarged opening came into direct contact with the surrounding water in which these animals lived. When the animal moved, the hair cells reacted to the flow of water that propagated into this cavity. The cavity developed a channel-like shape in order to be able to better pass on currents to the hair cells. This (possibly) resulted in the side-line system, which was present in almost all segments and enabled the development of an important sense of long-range location.
The active principle of the statocysts in the third segment underwent a fundamental change in the course of evolution, resulting in the development of the vestibulocerebellum. Only then did the line of vertebrates begin, whose neurological speciality is the possession of a cerebellum. In this respect, all vertebrates could also be called cerebellum animals.
Hint:
We assume
that there were other head segments that specialised in certain receptor types
or signal types. For example, there was certainly a head segment that served to
control the most elementary life processes and whose segment contained the later
hypothalamus of future vertebrates. The fact that visual signals were also
evaluated there is shown by the nucleus suprachiasmaticus, which acts as a
temporal clock. In this paper we will limit ourselves to a selection of
modalities.
Theorem of existence of a segment with a
second pair of eyes
Below the vestibular segment there was at least one secondary segment with another pair of eyes.
Many segmented animals have several segments that have eyes. It is possible that in the animals whose line led to the vertebrates, a fusion and reduction took place so that only one additional segment had a pair of eyes. This was necessarily arranged after the vestibular segment and had only two compound eyes, while the primary eyes of the second segment had to develop into lens eyes. The proof is given step by step in the following sections.
Hint:
The pairs of eyes arranged at different segments were included in the mean value systems of the respective segments. The latter often served other purposes in different segments, especially in the head segments. The secondary eyes of the sixth segment gave rise to the parietal eyes in the lower vertebrates, which were mainly used to determine the mean brightness. They were included in the mean value system of the hypothalamic region and provided, for example, the nucleus suprachiasmaticus, which functions as a circadian clock, with the necessary visual mean value signals. In higher vertebrates, a reduction was also carried out here, although the primary function was maintained.
In addition to a specialization of the segments in terms of sensory modalities, a further specialization was also observed. While initially all segments (except the head and tail segment) were in principle identical and had the same equipment of primitive organs such as heart or kidneys, this changed in the course of evolution. In the end there was only one heart, one stomach, two kidneys etc.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan