Vertebrate brain theory
ISBN
978-3-00-064888-5
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
3.9 The reconstruction of the vestibular sense and the development of the cerebellum
The vestibular sense underwent constructive changes in the course of evolution. In this monograph we refer to its earliest form as the paleo-oestibular sense.
The paleovestibular sense was mechanically and constructively not the best solution.
A disadvantage for the vestibular sense was its dependence on the accidental presence of a grain of sand in the vestibular cavity. The movements of the grain of sand also caused the hair cells to wear out. These disadvantages led to an evolutionary transformation of the vestibular sense.
A protective gelatinous layer was formed, in which the hair cells were embedded to protect them and prolong their life. The rolling grain of sand was replaced by stationary, small crystalline secretions (statoconia) on the gelatinous mass, which were formed by the living being itself. This is how the neovestibular sense was created.
The self-generated statoconies were now distributed more or less evenly over the jelly. Due to their own weight, they burdened the hair cells in the vicinity. However, the hair cells located at the deepest point of the vestibular cavity were only slightly bent and were therefore the least excited.
Hair cell theorem of the neovestibular sense
The hair cells of the neovestibular sense are comparable to firmly clamped elastic leaf springs, to whose upper end a small weight (the statokonie) is attached. Their angle of inclination depends on their location in the vestibular cavity. The more the leaf spring is inclined against the vertical, the greater the bending angle. This is converted by the hair cell into a sequence of action potentials. The larger the bending angle, the higher the rate of fire.
The vestibular cavity formed a cavity. The hair cell that was at the deepest point of the cavity was sheared the least from its normal position. The low weight of the statokonie caused only negligible bending.
All other hair cells were excited by the weight of the statoconies connected to them, the greater the deviation from the vertical, the more excited the hair cells were. This is comparable to the deflection of a leaf spring, which is loaded with a small weight at its upper end. Its deflection is smallest when it is exactly vertical, whereby the weight must not be too great. With increasing angle of inclination, the deflection becomes greater. The weight of the statokonie causes a considerable bending of the hair cell via the long lever arm when its orientation deviates more from the vertical.
We call this vestibular system, which has been converted in this way, neovestibular sense. The neovestibular sense was minimum coded. The hair cell with a minimum firing rate coded the actual angular deviation of the vestibular system from the vertical.
The reconstruction of the vestibular sense did not start suddenly, it will have developed successively over many generations. In the course of millions of years, further specialisations of the hair cells were added, which modified the signal evaluation.
We assume that during the remodelling of the vestibular sense all previous nerve cells and their axons were preserved and that the neuronal circuitry did not change significantly at first.
With the transition from the paleo- and neo-anestibular sense, the previous signal evaluation achieved the opposite of the desired position correction. The previous signal maximum had been replaced by a signal minimum during the conversion of the vestibular sense, and the maximum-coded paleo-ovestibular sense was thus converted to the minimum-coded neovestibular sense. The signal minimum was not sufficient for effective muscle control, because a muscle needs a strong excitation signal. A zero signal has no effect at all, and a very weak signal has almost none.
Theorem of coding types of the vestibular sense
The paleovestibular sense was maximum coded. The hair cell with the highest rate of fire was located at the lowest point of the vestibular cavity. Its output signal - with the interposition of the contralateral signal reflection in the crossing floor - controlled the motoneurons of the muscles, which initiated a corrective movement.
The neovestibular sense was minimum coded. The hair cell with the lowest rate of fire was located at the deepest point of the vestibular cavity. However, it was not able to generate muscle movement, the output signal was too weak for that. The output signals of the remaining hair cells were stronger, but their axons - with the interposition of other projection neurons - moved to motoneurons, the activation of which increased the deviation from normal position. The only hair cell that was connected to the motoneuron of the correct muscle via axons and intermediate neurons remained unexcited in the neovestibular sense instead of firing at maximum as before.
In the following figure you first see the minimum coded output of the neovestibular sense. The minimum encodes the inclination angle of the living being. If the body posture changes, the minimum moves to a new position:
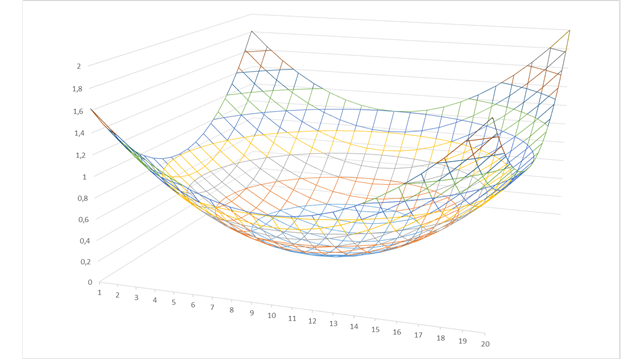
Figure 12 - Neural excitation of the minimum coded vestibular sense
Diese Codierungsform ist unbrauchbar. Spiegelt man die Funktion dagegen an einer waagerechten Ebene, die etwa in der Höhe des Mittelwertes der Funktion verläuft, entsteht eine brauchbare Signalversion, wie nachfolgende Abbildung zeigt. Eine solche Transformation bezeichnen wir als Signalinversion .
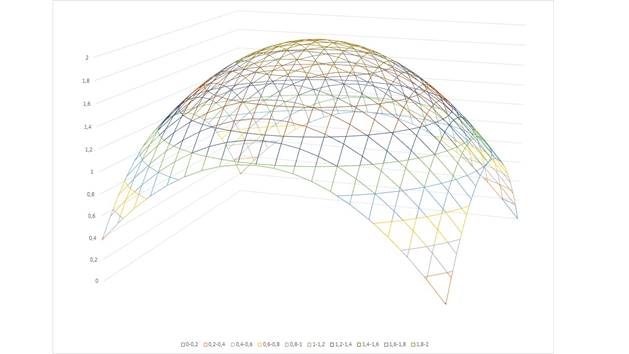
Figure 13-
Inverted output of the neovestibular sense -maximum coded
The useless signal minimum becomes a usable signal maximum by inversion. Nature proved to be able to realize such a signal inversion by means of a special neural circuit. We call such a circuit an inversion circuit.
The necessity to transform the minimum coded signals of the neovestibular sense into a maximum coded form led to the development of the vestibulocerebellum and the lateral vestibular nucleus. This is shown in the following chapter.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan