Vertebrate brain theory
ISBN
978-3-00-064888-5
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
4 The effects of signal divergence in the cerebellum system
The original spinocerebellum belongs to the contralateral subsystem, the ipsilateral signals reach the opposite side via the nucleus ruber and the nucleus olivaris. In this chapter we analyze the consequences of redundancy in this system.
Redundancy occurs when signals are divergently distributed over several, often even many signaling pathways before reaching their target structure, so that the failure of individual signaling pathways does not have any adverse consequences. Of course, these signals must be combined in a convergence structure before reaching their target structure.
Both signal divergence and subsequent signal convergence are subject to certain laws of nature, since they are usually realized in neuron nuclei or neuron layers that belong to the grey matter of the nervous system. The absence of myelin in these structures causes a distance-dependent attenuation of the signals, which leads to the occurrence of extremely interesting and important phenomena. This monograph shows that vertebrates owe their intelligence to precisely these phenomena. This chapter examines the effects of redundancy in the contralateral system, whose most prominent structure is the spinocerebellum. In the older literature this system is also called extrapyramidal system.
4.1 Computing with neural signals
Up to now we have used the term fire rate in a popular way. It was clear to us that the fire rate represents a signal strength and that the fire rate increases with the signal strength. But now we will try to be more precise. If we want to show that the vertebrate brain calculates with signals, that there are different signal forms, and that these are transformed into each other by mathematical transformations, then we must also interpret the concept of fire rate mathematically.
Fire rates are primarily caused by the activities of receptors acting on associated ganglion cells. We now look at the relationship between signal strength and fire rate.
Definition: greatness, original signal
A measurable variable u, whose measured values are measured in a measuring
interval
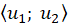 we call it greatnesswhen
receptors transform
this size into a rate of fire. We assign a
modality of its own to the quantity u. Each concrete value of the quantity u is
called the primal signal. The rate of fire belonging to this value is called the
neuronal signal of the quantity, the function f = f(u) is the
signal function of the quantity u.
we call it greatnesswhen
receptors transform
this size into a rate of fire. We assign a
modality of its own to the quantity u. Each concrete value of the quantity u is
called the primal signal. The rate of fire belonging to this value is called the
neuronal signal of the quantity, the function f = f(u) is the
signal function of the quantity u.
Most of the basic variables are continuous functions. We therefore restrict our consideration to this class of functions without constantly pointing it out.
Definition On-Signal, Off-Signal
If the continuous signal function f = f(u) is strictly monotonically growing, we call the signal on-signal. If it is strictly monotonically decreasing, then there is an off-signal.
Theorem of the exponential relationship between magnitude and neuronal signal in motor function
The signal function f = f(u), which describes the relationship between the signal strength of a continuous original quantity and the fire rate f in the motor function derived from it, is (approximately) exponential. If f1 is the rate of fire at the left end of the interval, i.e. f1 = f (u1), and f2 is the rate of fire at the right end of the interval, i.e. f2 = f (u2), the equation
![]()
and for off-signals
![]() ,
,
where k is an independent constant that can take on a different value for each modality.
A signal function can be interpreted as the mapping of a primal signal interval <u1, u2> to a fire rate interval <f1, f2>.
Theorem of mapping a primal signal interval to a fire rate interval
Each exponential on-signal function produces a unique and continuous
representation of the original signal interval
![]() with
with
![]() and
and
![]()
to a fire rate interval
![]() according to
according to
![]() and
and
![]() .
.
Each exponential off-signal function produces a unique and continuous
representation of the original signal interval
![]() to a fire rate interval
to a fire rate interval
![]() according to
according to
![]() and
and
![]() .
.
This unambiguous mapping of both intervals makes it relatively easy for us to draw conclusions about the greatness of the fire rate. For example, if the original magnitude is the light frequency in a frequency interval, the rate of fire ultimately represents a color, because each light frequency of visible light can be uniquely assigned a color. The more detailed background of color vision will be described at the end of this chapter.
neural mean theorem
If an on-signal function f= f(u) is in the interval
![]() defines and
defines and
![]() and is
and is
![]() the corresponding signal average value in the interval, then
the corresponding signal average value in the interval, then
![]() .
(Valid for On
functions)
.
(Valid for On
functions)
The following applies analogously to an off-signal function
![]() .
(Valid for off functions)
.
(Valid for off functions)
The importance of the fire rate mean value will become particularly clear in this chapter. The relationship between a signal and its inverted signal is also interesting.
The maximum achievable fire rate of projection neurons is limited upwards, we can consider it as a constant. Then (almost) all modalities are compatible with each other.
Theorem of compatibility of modalities with strictly monotone signal functions
The intervals of the different primal sizes with strictly monotone signal functions, to which different modalities correspond, all map the primal intervals to the same fire rate interval <0; fmax>, where fmax represents the maximum achievable fire rate of the involved projection neurons and is a kind of technical constant.
Conversely, the value of any primal quantity could be assigned to any fire rate, if only one suitable type of receptor exists which transforms this primal quantity into a fire rate. For signal functions that are not strictly monotonous, this is of course not true. If the original quantity is discontinuous or even discrete, these statements are not true either.
Theorem of the quotient of a signal and its inverted signal
From the equation
![]() according to (18.3.4) follows for the relationship between a signal and
its inverted signal
according to (18.3.4) follows for the relationship between a signal and
its inverted signal
the equation
![]() .
(2.1.3)
.
(2.1.3)
Many types of receptors have a composite characteristic curve, which in part rises in a strictly monotonous manner until it reaches a resonance point where the rate of fire becomes maximum, only to fall again in a strictly monotonous manner when the original size is further increased. We call such receptors resonance point receptors. Their characteristic curve resembles a Gaussian bell curve and is approximately described by a combination of two exponential functions. For example, the rate of fire of those retinal ganglion cells that form the signal red increases with the left-sided approximation of the light frequency to the frequency of red light, but decreases again with a further increase of the light frequency. The resonance maximum is thus reached at red, and the firing rate decreases equally to the left and to the right when there is a deviation. Therefore there is no clear correlation between light frequency and firing rate. Nature solved this ambiguity problem by developing alternative visual dyes, which, for example, have a resonance maximum at other light frequencies, such as green or blue. This will be analyzed later in this monograph.
Theorem of the fire rate of resonance point receptors
If um the resonance point of a receptor, the signal function can be (approximately) described by the equation
![]() (2.1.6)
(2.1.6)
The Equation
![]() (2.1.7)
(2.1.7)
Enables the calculation of the original value from the function values of the signal function.
Instead of the great magnitude u, the amount |u| of the great magnitude is inserted into the equations. The signal function thus becomes symmetrical um the center point at which the function takes on the value fm.
Examples of such receptors are the thermoreceptors or the rods for color vision. For receptors with a resonance point, the clear projection of the original value to the signal value is omitted. The same firing rate may mean different original values here, since original values located symmetrically to the centre around the centre always provide the same firing rate.
In order to eliminate this ambiguity, a special type of signal transformation has become established which exploits the ability of the spinocerebellum to invert signal values.
This is explained in more detail below.
In this monograph, a relationship is established between the exponential sensitivity characteristic of receptors and the exponential attenuation of neuronal excitations that propagate to non-marrow axons. The latter is described by the cable equation for non-markless fibers.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan