Theorie der neuronalen Schaltung des Gehirns
und des analytischen Denkens
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
Teil 1.2. Typisierung von neuronalen Kerngebieten
Für die nachfolgenden Erklärungen benötigen wir einige elementare Grundbegriffe, die nun definiert werden.
Definition 1.3: Kerngebiete
Ansammlungen von Neuronen werden als Kerngebiete bezeichnet.
Kerngebiete können nach der Anzahl der Input- und Outputleitungen unterschieden werden.
Definition 1.4: Umschaltkerne
Die Anzahl der Inputleitungen ist etwa gleich der Anzahl der Outputleitungen, jedoch benutzen Input und Output meist unterschiedliche Transmitter, so dass von einem Transmitter auf einen anderen umgeschaltet wird.
Definition 1.5: Konvergenzkerne
Die Anzahl der Inputleitungen ist deutlich größer als die Anzahl der Outputleitungen.
Definition 1.6: Divergenzkerne bzw. Divergenzschicht
Die Anzahl der Inputleitungen ist deutlich kleiner als die Anzahl der Outputleitungen.
Unter Input- bzw. Outputleitungen mögen wir uns die Axone der Nervenzellen vorstellen, die der Signalweiterleitung dienen. Die Begriffe Umschaltkern, Konvergenzkern und Divergenzkern nutzen wir zur Klassifizierung der neuronalen Kerne.
Theorem 1.2: Die Substantia nigra pars compacta ist ein Umschaltkern für den Cortexoutput
Die Substantia nigra pars compacta erhält einerseits den Output der Clusterneuronen und andererseits den Output der Aktivitätsneuronen. Beide Arten von Output sind glutamaterg und werden auf den Transmitter Dopamin umgeschaltet. Während die Clusterneuronen auf Dopamin vom Subtyp D2 umgeschaltet werden und ihre Zielneuronen in der Matrix des Striatums hemmen, werden die Aktivitätsneuronen der Cortexcluster auf Dopamin vom Subtyp D1 umgeschaltet und wirken auf ihre Zielneuronen in den Striosomen des Striatums erregend.
Die Substantia nigra pars compacta ist ein neuronaler Kern des Basalgangliensystems. Das Theorem 1.2 unterstellt, dass es die Hauptaufgabe der Substantia nigra pars compacta wäre, den dopaminergen Input vom Cortex auf den Transmitter Dopamin umzuschalten. Dies macht Sinn, weil der Output dieses Kernes wieder in Richtung des Cortex geschickt wird, quasi in die Richtung, aus der er ursprünglich kam. Weil er aber vor dem Richtungswechsel auf einen anderen Transmitter umgeschaltet worden ist, kann das neuronale System Cortexoutput und nigralen Output gut auseinanderhalten.
Theorem 1.2 unterstellt jedoch auch (eine bereits bekannte) Spezifizierung der Neuronen der Substantia nigra pars compacta in zwei Gruppen. Eine Gruppe nutzt im Zielgebiet, dem Striatum, die D1-Rezeptoren, die vom Transmitter Dopamin erregt werden. Diese Gruppe werden wir als dopaminerge Neuronen vom Subtyp D1 bezeichnen. Die andere Neuronengruppe des Kerngebietes nutzt im Striatum ebenfalls dopaminerge, aber hemmende D2-Rezeptoren. Diese Gruppe von Neuronen werden wir als dopaminerge Neuronen vom Subtyp D2 bezeichnen.
Das Theorem 1.2 unterstellt eine Ursache für die bekannte Kompartimentierung des Striatums in die Matrix und in die Striosomen. Bekannt ist (im Jahre 2011), dass die dopaminergen Axone aus der Substantia nigra pars compacta einerseits vermittels D1-Rezeptoren erregend und andererseits über D2-Rezeptoren hemmend wirken.
Das Theorem 1.2 unterstellt einen bis heute noch nicht erkannten, neuen Sachverhalt dieser Kompartimentierung. Es wird behauptet, dass die Aktivitätsneuronen aller Cluster der Cortexrinde in der Substantia nigra pars compacta auf dopaminerge Neuronen vom Subtyp D1 umgeschaltet werden und anschließend die Neuronen der Striosomen des Striatums erregen, während die Signalneuronen aller Cortexcluster in der Substantia nigra pars compacta auf dopaminerge Neuronen vom Subtyp D2 umgeschaltet werden, um anschließend die Neuronen der Matrix des Striatums zu hemmen.
Der tiefere Sinn einer solchen komplementären Projektion und der Kompartimentierung des Striatums in Matrix und Striosomen wird in dieser Theorie erstmals umfassend aufgeklärt.
Skizze 1.2: Substantia nigra als Umschaltkern von Glutamat auf Dopamin
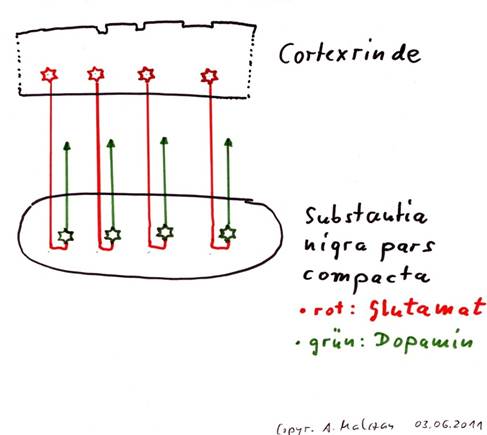
Umschaltkerne erfüllen im Gehirn eine sehr wichtige Aufgabe, obwohl sie ein Signal nur von einem Transmitter auf einen anderen Transmitter umschalten. Hierdurch wird es jedoch möglich, Input und Output auseinanderzuhalten. Würde das gesamte Nervensystem nur eine Transmitterart verwenden, so wäre, da jedes Neuron mit jedem Neuron transmitterkompatibel wäre, ein totales Signalchaos die Folge. Erst die Verwendung unterschiedlicher Transmitter ermöglicht es, dass ein Neuron aus einem riesigen Signalangebot nur die Axone anzapft, zu denen es transmitterkompatibel ist.
Theorem 1.3: Das Striosomensystem des Striatums ist ein Umschaltkern
Die Aktivitätsneuronen (Mittelwertneuronen) der Cortexcluster projizieren erregend in den Umschaltkern Substantia nigra pars compacta, wo sie auf Dopamin vom Subtyp D1 umgeschaltet werden. Dieser Output wird in den Striosomen des Striatums auf den Transmitter GABA umgeschaltet. Daher ist das Striosomensystem ein Umschaltkern.
Es wird sich noch zeigen, dass das Matrixsystem des Striatums andere Funktionen hat.
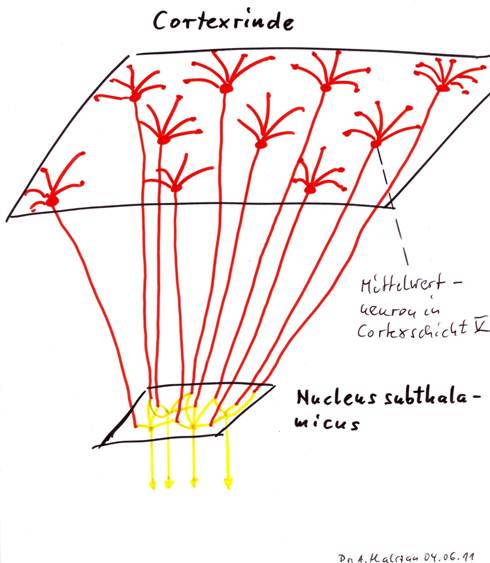
Theorem 1.4: Der Nucleus subthalamicus ist ein neuronaler Konvergenzkern
Der Nucleus subthalamicus empfängt den Output der Aktivitätsneuronen der Cortexcluster. Seine Neuronen bilden aus den gelieferten mittleren Signalpegeln durch erneute Mittelwertbildung Gebietsmittelwerte, die den Aktivitätszustand größerer Cortexregionen repräsentieren, die selbst aus vielen Cortexclustern bestehen.
Insofern sind die Outputneuronen des Nucleus subthalamicus Aktivitätsneuronen zweiter Stufe. Ihr Output ist meist ein mehr oder minder starkes Dauersignal, dessen Feuerrate die Gebietsaktivität des zugehörigen Cortexgebietes repräsentiert.
Beachtet werden muss auch, dass die Cortexrinde oft komplementäre Signale empfängt, also solche, bei denen es sowohl eine On-Variante als auch eine Off-Variante gibt (z. B. im visuellen primären Cortex). Dadurch gibt es bei den Aktivitätsneuron des Nucleus subthalamicus fast immer eine Inputzufuhr. Nur bei völliger Signallosigkeit großer Cortexgebiete könnten die Outputneuronen des Subthalamicus ebenfalls schweigen.
Skizze 1.3: Der Nucleus subthalamicus als Konvergenzkern für den Cortexoutput aus Schicht V
Skizze 1.4: Konvergenz und Divergenz als Grundprinzipien des Gehirns
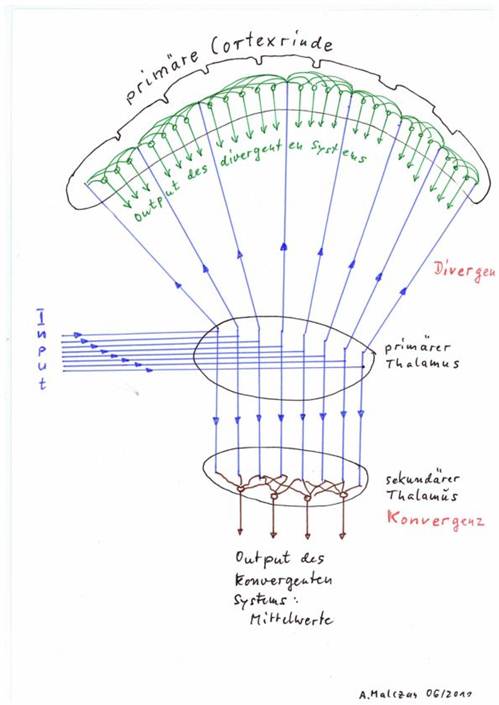
Konvergenzkerne und Divergenzkerne können natürlich zusätzlich auch Umschaltkerne sein. Hingewiesen werden muss darauf, dass letztendlich die Rezeptoren der Ionenkanäle darüber entscheiden, ob ein Transmitter erregend oder hemmend wirkt. Zu vielen Transmittern gibt es durchaus mehrere Typen von Rezeptoren mit unterschiedlicher Wirkung.
In dieser Theorie soll auch gezeigt werden, dass das Gehirn computerähnlich arbeitet. Daher ist es erforderlich, einige elementare Grundbegriffe aus der Computerwelt einzuführen.
Definition 1.7: Nullsignal
Einem Neuron, welches keine Aktionspotentiale erzeugt, ordnen wir die Feuerrate f = 0 zu und bezeichnen seinen Output als Nullsignal. Es besitzt dann den Signalwert Null.
Definition 1.8: Einssignal
Einem Neuron, welches Aktionspotentiale mit einer beliebigen Feuerrate f größer als Null erzeugt, ordnen wir das Einssignal zu. Es besitzt den Signalwert 1.
Somit hat ein tatsächlich feuerndes Neuron den Signalwert 1, während ein nicht feuerndes den Signalwert Null besitzt. Damit repräsentieren Neuronen nach der „Alles-oder-Nichts-Regel“ einen binären Signalwert. Dies ist der erste Hinweis auf computerähnliches Verhalten.
Skizze 1.5: Neuronales Einssignal – nicht unbedingt frequenzstabil:

Neurologen pflegen Nervenzellansammlungen als Kerne zu bezeichnen. Der Nucleus subthalamicus ist ein solcher Kern. Seine Outputaxone führen (meist) ein Dauersignal, welches durch doppelte Mittelwertbildung über der Clustermenge des Cortex entsteht. Da aber Dauersignale durch tatsächlich feuernde Neuronen erzeugt werden und deren (binärer) Signalwert der Wert 1 ist, erzeugen die Outputneuronen des Nucleus subthalamicus letztendlich Einssignale. Diese haben jedoch meist unterschiedlich starke Feuerraten.
Definition 1.9: Positiver Einssignalkern
Erzeugt ein Neuronenkern aus den ihm zugeführten oder durch ihn hindurchziehenden erregenden Mittelwertsignalen erregende Dauersignale (Einssignale), so wird er als positiver Einssignalkern bezeichnet.
Definition 1.10: Negativer Einssignalkern
Erzeugt ein Neuronenkern aus den ihm zugeführten oder durch ihn hindurchziehenden erregenden Mittelwertsignalen hemmende Dauersignale (Einssignale), so wird er als negativer Einssignalkern bezeichnet.
Theorem 1.5: Der Nucleus subthalamicus ist ein positiver Einssignalkern
Der Nucleus subthalamicus ist ein positiver Einssignalkern. Seine Outputneuronen erzeugen in den meisten Fällen erregende Einssignale.
Skizze 1.6: Entstehung von Einssignalen (Dauersignalen) aus einer ununterbrochenen Folge von Aktionspotentialen

Veränderte Zeitauflösung:
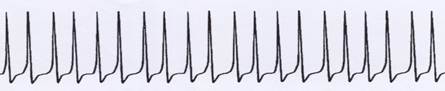
Noch mehr zeitlich gestaucht:
Zeitlich sehr eng gestaucht: Beispiel für ein Einssignal (Dauersignal)





Wenn jedoch die Cortexaktivität in besonderen Fällen (Schlaf, tiefe Ohnmacht, Koma) in bestimmten, größeren Regionen stark vermindert ist, kann ein Teil des Outputs des Nucleus subthalamicus auch das Nullsignal sein.
Neurologen mögen sich wundern, dass ein neuronales Dauersignal als „Einssignal“ bezeichnet wird, ein nicht existierendes Signal dagegen als „Nullsignal“. Aber eine neuronale Zahlentheorie benötigt auch die Grundelemente der Zahlentheorie. Das Nullelement und das Einselement sind die Grundlage aller Zahlenräume.
Weiterhin sind Null und Eins als numerische Entsprechungen der logischen Werte „Wahr“ bzw. „Falsch“ wichtige Grundelemente der Aussagenlogik und ermöglichen binäres Rechnen.
- ← Teil 1.1. Cortexcluster, Aktivitätsneuron und mittlerer Signalpegel
- Index
- Teil 1.3. Die einfache und die doppelte Negation neuronaler Signale →
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan