Theorie der neuronalen Schaltung des Gehirns
und des analytischen Denkens
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
Teil 1.3. Die einfache und die doppelte Negation neuronaler Signale
Computer rechnen mit digitalen Signalen. Bisher haben wir erkannt, dass der Output von Neuronen als digitales Signal aufgefasst werden kann. Entweder das Neuron feuert und hat den Signalwert 1, oder es feuert nicht und hat den Signalwert Null.
Doch können Neuronen mit diesen Signalwerten auch binäre Operationen ausführen, so wie moderne Computer dies tun?
Hier sollten wir den Begriff der Hüllkurve einführen. Jedem Outputsignal eines Neurons, dessen Output sich zeitlich beliebig ändern kann, kann man eine Hüllkurve zuordnen, wenn man die Aktionspotentiale als Zeitfunktion darstellt. Diese Hüllkurve verläuft, abgesehen von Sprungstellen, immer parallel zur t-Achse (Zeitachse) und hat nur zwei mögliche Werte: 0 oder 1.
Skizze 1.7: Hüllkurve eines neuronalen Signals und Binärwert des Signals
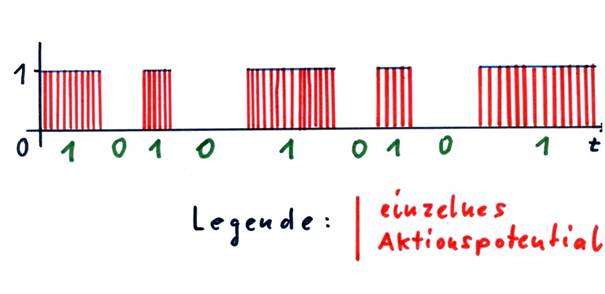
Nervenzellen stehen als -Rechenoperationen- nur die Erregung oder die Hemmung zur Verfügung. Die Existenz von Einssignalen im Nucleus subthalamicus ist bereits nachgewiesen worden.
Wir untersuchen nunmehr, was passiert, wenn ein Neuron N1 ein Signal s1 erzeugt und dieses Signal ein vorhandenes Dauersignal hemmt. Die einfachste Art der Hemmung ist die vollständige Hemmung.
Skizze 1.8: Hemmung eines Eissignals durch ein Signal
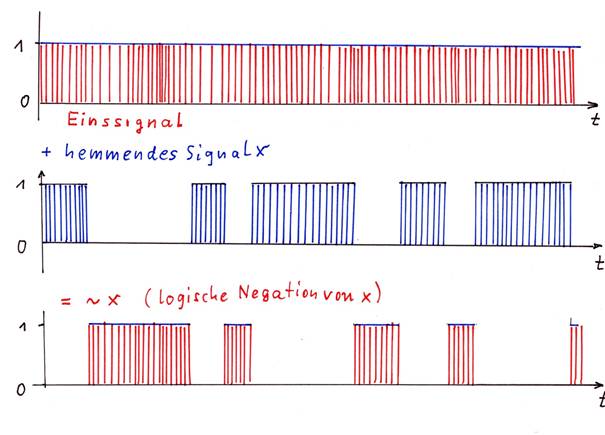
Das Einssignal (rot) wird von Signal x (blau) total gehemmt, das Ergibnis ist das Signal ~x (rot), welches die Negation des Signals x darstellt.
Hier wird deutlich: Hemmt ein Signal S ein Einssignal vollständig, so entsteht als Ergebnis die logische Negation ~S des Ausgangssignals S.
Theorem 1.6: Negationstheorem
Hemmt ein neuronales Signal S ein Dauersignal D = 1 stark und vollständig, so entsteht als Output das negierte Signal ~S.
Die Negation ist jedoch eine elementare binäre Operation. Somit ist bewiesen worden: Das Gehirn kann wenigstens eine binäre Operation ausführen, nämlich die Negation eines Signals. Dazu benötigt es jedoch ein Dauersignal.
Man beachte, dass hier eine vollständige Hemmung vorliegen muss. Bei einer relativen, vielleicht sogar schwachen Hemmung bleibt immer dort ein Restsignal übrig, wo bei vollständiger Hemmung das Nullsignal vorliegen würde.
Bereits zu Beginn des Abschnitts wurden neuronale Kerne definiert. Hier können zwei neue neuronale Kerne definiert werden.
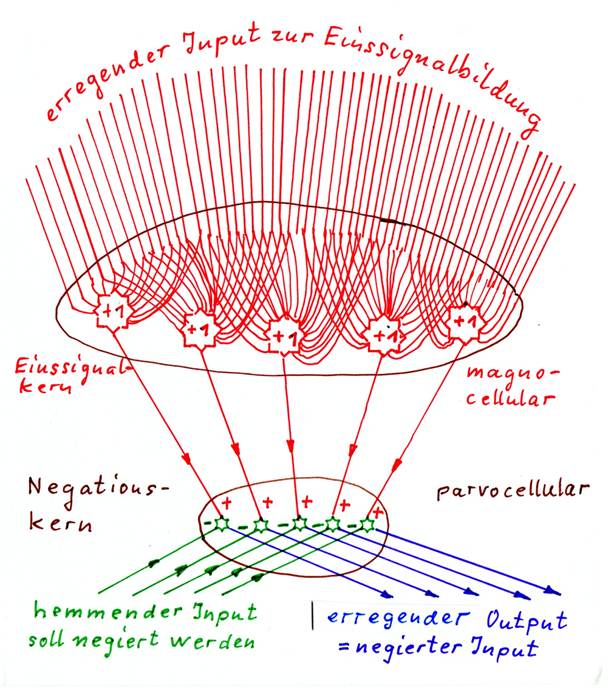
Definition 1.11: Positiver Negationskern
Erhält ein Kern einen erregenden Input von einem Einssignalkern und einen hemmenden Input von einem Umschaltkern und erzeugt durch Hemmung der zugeführten Einssignale zu jedem hemmenden Input die Negation als Output, so ist dieser Kern ein positiver Negationskern, sein Output ist erregend.
Ein Negationskern kann jedoch auch hemmenden Output liefern.
Definition 1.12: Negativer Negationskern
Erhält ein Kern einen hemmenden Input von einem Einssignalkern und einen hemmenden Input von einem Umschaltkern und erzeugt durch Hemmung der zugeführten Einssignale zu jedem hemmenden Input die Negation als Output, so ist dieser Kern ein negativer Negationskern, sein Output ist hemmend.
Theorem 1.7: Der Globus pallidus interna ist ein negativer Negationskern für die Striosomen
Der Globus pallidus interna erhält GABAergen Input aus den Striosomen des Striatums und erregenden Einssignalinput vom Nucleus subthalamicus. Sein Output besteht aus dem negierten Striosomenoutput und wirkt hemmend auf die Zielneuronen.
Festlegung: Neuronen, die ein erregendes Einssignal erhalten, kennzeichnen wir in Skizzen durch das Symbol "+1". Neuronen, die ein hemmendes Einssignal als Input erhalten, kennzeichnen wir in Skizzen durch das Symbol "-1".
Skizze 1.9: Der Globus pallidus interna als negativer Negationskern für die Striosomensignale
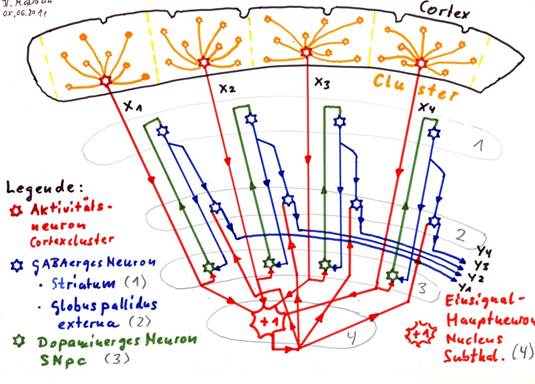
Korrektur: Beschriftung: Globus pallidus interna (2) [statt externa!]
Erläuterung zur Skizze 1.9:
In jedem Cortexcluster erzeugt das Aktivitätsneuron ein Mittelwertsignal, in der Skizze 1.9 als x1, x2, x3 und x4 gekennzeichnet.
Diese Aktivitätsneuronen projizieren einerseits alle in den Nucleus subthalamicus (4), der daraus Mittelwerte zweiter Stufe erzeugt, die (meist) daueraktive Einssignale sind (rot +1).
Gleichzeitig wird jedes Mittelwertsignal im Umschaltkern Substantia nigra pars compacta (3) von Glutamat (rot) auf Dopamin (grün) (Subtyp D1 - erregend) umgeschaltet. Die erregenden dopaminergen Signale erreichen das Striatum (genauer die Striosomen im Striatum) (1) und enden auf GABAergen Neuronen, die diese Signale von Dopamin (grün) auf GABA (blau) umschalten. Die Axone der GABAergen Striatumneuronen verzweigen sich und projizieren einerseits hemmend zur Substantia nigra pars compacta (3) als auch zum Globus pallidus interna (2). Die Neuronen des Globus pallidus interna erhalten zwei Signaleingänge: Erstens erhält ein jedes Neuron ein glutamaterges Einssignal aus dem Nucleus subthalamicus (rot +1). Dadurch wären diese Outputneuronen dauererregt und würden ein GABAerges Dauersignal (Einsignal) aussenden, wenn nicht als zweiter Input die Hemmung durch den GABAergen Input der striatalen Neuronen (blau) einwirken würde.
Der Output y1, y2, y3, y4 in der Skizze ist also der negierte Cortexoutput, also die negierten Mittelwertsignale x1, x2, x3 und x4. Dieser Output ist hemmend. Außerdem ist der Output - verglichen mit dem Originaloutput des Cortex - leicht zeitverzögert, da die Signale einen relativ weiten Weg zurücklegen mussten.
Wäre der Output erregend statt hemmend, läge hier ein positiver Negationskern vor.
Der Einfluss der hemmenden Projektion der Striosomenneuronen des Striatums (2) auf die Substantia nigra pars compacta (4) wird später erörtert werden. Diese Projektion ermöglicht es, das Basalgangliensystem als Schwingkreis aufzufassen, der den Systemtakt des Gehirncomputers erzeugt. Diesem Umstand ist ein eigenes Kapitel gewidmet.
Nicht immer werden die nötigen (positiven oder negativen) Einssignale einem Negationskern von außen zugeführt. Beim Globus pallidus interna ist dies jedoch der Fall.
Wenn die Einssignale nicht von externen Kernen kommen, müssen sie im Negationskern selbst erzeugt werden. Dazu benötigt ein Negationskern dann zusätzlichen, erregenden Input von magnocellularen Neuronen, am besten von Mittelwertneuronen.
Aus ihren Mittelwertsignalen entstehen durch erneute Mittelwertbildung innerhalb des Negationskerns die zur Negation nötigen Einssignale. Daher können wir die Typen der Negationskerne erweitern.
Definition 1.13: Positiver Negationskern mit interner Einssignalbildung
Ein positiver Negationskern mit interner Einssignalbildung erhält erregenden magnocellularen Input und bildet daraus erregende Einssignale. Eine weitere hemmende Inputart hemmt die Einssignale und wird somit an ihnen negiert. Der negierte Output des Kerns ist erregend.
Definition 1.14: Negativer Negationskern mit interner Einssignalbildung
Ein negativer Negationskern mit interner Einssignalbildung erhält erregenden magnocellularen Input und bildet daraus hemmende Einssignale. Eine weitere hemmende Inputart hemmt die Einssignale und wird somit an ihnen negiert. Der negierte Output des Kerns ist hemmend.
Negationskerne mit interner Einssignalbildung müssen also immer ein eigenes Einssignal bilden. Dazu benötigen sie magnocellulare Mittelwertneuronen, die erregenden oder hemmenden Einssignalinput bilden. Diese magnocellularen Neuronen mit riesigen Dendritenbäumen sind ein verräterisches Kennzeichnen der Negationskerne mit interner Einssignalbildung. Immer erhalten sie erregenden Input, obwohl sie selbst erregend oder hemmend wirken können.
Der zu negierende Input muss hemmend sein, da er das gebildete Einssignal zu unterdrücken hat. Daher ist ein hemmender Input ein weiteres Kennzeichen solcher Kerne. Deshalb müssen erregende Signale vor einer Negation in einem vorgelagerten Umschaltkern auf hemmende Transmitter umgeschaltet werden.
Erkennungsmerkmale für Negationskerne mit innerer Einssignalbildung
-Erregender Input auf große, magnocellulare, integrativ wirkende Neuronen, die das Einssignal bilden (Einssignalneuronen, Hauptneuronen). Diese Neuronen sind an ihren riesigen Dendritenbäumen zu erkennen. Sie können erregend oder hemmend wirken.
-Vorzugsweise sollte der Einssignalinput bereits von großen, magnocellularen Mittelwertneuronen stammen, z. B. aus der Schicht V des Cortex, damit tatsächlich ein stabiles Dauersignal (Einssignal) gebildet wird und kein nur zeitweilig aktives Mittelwertsignal
-Hemmender Input, der auf die Einssignalneuronen (Hauptneuronen) stark hemmend einwirkt (Synapse in der Nähe des Axonhügels bzw. zellkörpernah am Dendritenstamm).
-Der erregende Input für die Einssignalneuronen kann im Kern einfach an den Hauptneuronen enden (z. B. Striatum, genauer Matrix). Er kann aber auch durch diesen Kern hindurch geleitet werden und ihn wieder verlassen (z. B. Nucleus ruber).
-Negative Negationskerne mit innerer Einssignalbildung bestehen aus nur einem Kern, dessen magnocellulare Hauptneuronen hemmend wirken. Sie werden meist durch die durch diesen Kern hindurchziehenden, erregenden Axone übergeordneter Subsysteme erregt. Die gebildete Einssignalerregung wird von einer weiteren, hemmenden Inputart stark (total) gehemmt, wodurch letzterer Input negiert wird (z. B. Matrix des Striatums).
-Positive Negationskerne mit innerer Einssignalbildung bestehen aus zwei gut trennbaren Teilkernen: ein magnocellularer Kern erzeugt mit den riesigen Hauptneuronen die nötigen Einssignale, der parvocellulare Teilkern enthält die eigentlichen Negationsneuronen (z.B. Nucleus ruber, evtl. einige Thalamuskerne).
-Der erregende Input für die Einssignalbildung kann den Kern durchqueren, während er von den Hautpneuronen angezapft wird. Dies ist sehr oft tatsächlich der Fall. Aber ebenso könnte der erregende Input im Kern an den Hauptneuronen enden, ohne danach den Kern zu verlassen.
Erkennungsmerkmale für Negationskerne mit externer Einssignalbildung
-Der erregende Einssignalinput wird von einem externen Einssignalkern bereitgestellt.
-Der zu negierende Input muss immer hemmend sein. Notfalls muss er in einem externen Umschaltkern vom einem erregenden auf einen hemmenden Transmitter umgeschaltet werden.
-Jedes Outputneuron erhält zellkernnah, am wirksamsten direkt am Axonhügel sowohl den erregenden, fremderzeugten Einssignalinput als auch den stark hemmenden zu negierenden Input. Daher enthält der Kern meist nur kleinzellige, also parvocellulare Neuronen.
Im Hinblick auf die folgenden Kapitel sollte darauf hingewiesen werden, dass es neben den neuronalen Negationskernen eine weitere Kernklasse gibt, deren Erkennungsmerkmale denen der Negationskerne fast völlig gleichen. Es sind dies die Inversionskerne, die später beschrieben werden. Sie unterscheiden sich von den Negationskernen nur in einigen unauffälligen, aber funktionsbestimmenden Details.
Skizze 1.10: Negativer Negationskern - prinzipieller Aufbau
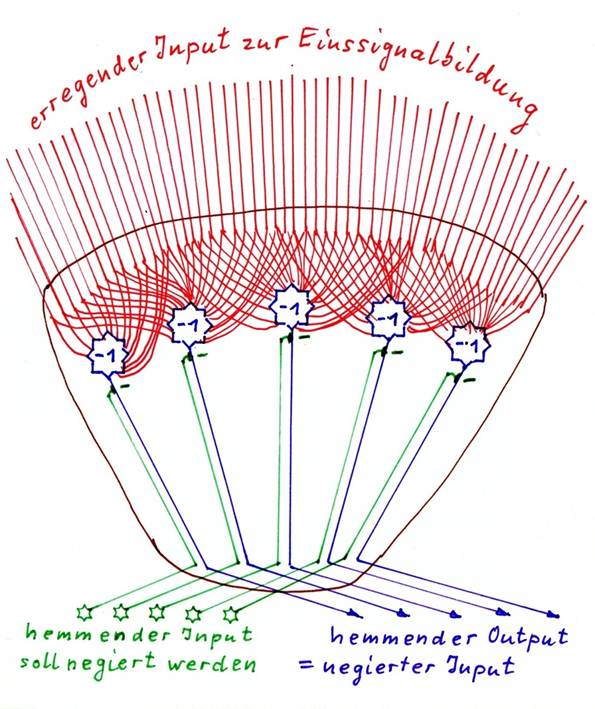
Skizze 1.11: Positiver Negationskern - prinzipieller Aufbau
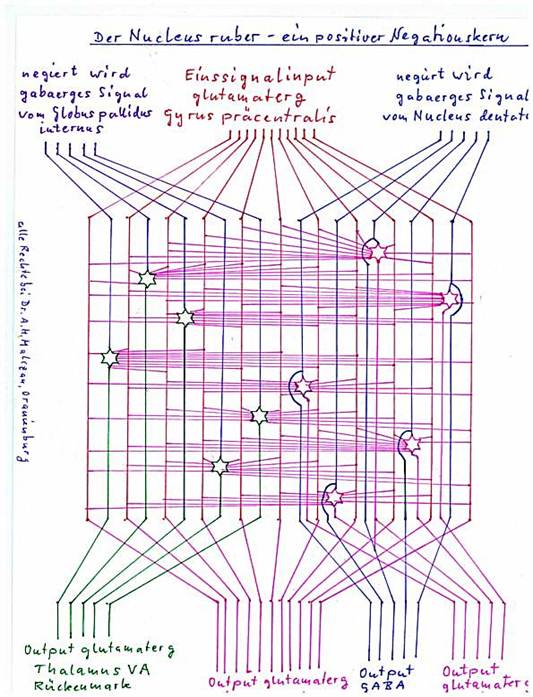
Theorem 1.8: Der Nucleus ruber ist für die Signale des Globus pallidus interna ein positiver Negationskern mit interner Einssignalbildung
Der Nucleus ruber erhält mehrere Arten von Input.
Vom Cortex erhält er erregenden Input. Bezogen auf die Cortexcluster erhält er sowohl den Output der Signalneuronen als auch den Output der magnocellularen Aktivitätsneuronen (Mittelwertneuronen). Letzterer Input wird im Nucleus ruber zur Bildung des positiven Einssignals herangezogen. Er stammt vorwiegend aus dem Gyrus präcentralis.
Der zu negierende Input stammt aus zwei Quellen. Zunächst liefert der Globus pallidus interna einen hemmenden, GABAergen Input, der die positiven Einssignale hemmt und an ihnen negiert wird. Somit verlässt der negierte Output des Globus pallidus interna den Nucleus ruber als erregendes Signal (in Richtung Nucleus olivaris inferior).
Die erregenden Signale aus dem Cortex, die der Bildung des positiven Einssignals dienen, werden zwar zur Einsignalbildung herangezogen, durchqueren jedoch den Nucleus ruber unbeschadet in Richtung Rückenmark.
Skizze 1.12: Der Nucleus ruber als positiver Negationskern
Es mag der Anschein entstehen, ein Großteil der neuronalen Kerne des Gehirns diene der Ne-gation von Signalen. Auffällig ist, dass solche Negationskerne oft hintereinander geschaltet werden. So wird der Striosomenoutput vom Globus pallidus interna negiert. Genau dieser Output erreicht den Nucleus ruber, um dort ebenfalls negiert zu werden. Doppelte Negation erzeugt in der Aussagenlogik aber das ursprüngliche Originalsignal. Dies ist bei der doppelten neuronalen Negation nicht mehr ganz der Fall.
Einerseits wird eine Frequenztransformation durchgeführt. Das Outputsignal hat in der Phase, wo es den Signalwert 1 annimmt, eine andere Frequenz als das Ursprungssignal. Die Outputfrequenz wird von der Frequenz des Einssignals bestimmt, welches durch das zu negierende Signal stellenweise unterdrückt wird.
Andererseits ergibt sich durch die Leitungslänge der beteiligten Axone eine Zeitverzögerung, da sich Aktionspotentiale nur mit endlicher Geschwindigkeit ausbreiten. Selbst eine Geschwindigkeit von 1 Meter pro Sekunde (1 m/s) ergibt pro Millimeter eine Zeitverzögerung von einer Millisekunde.
Eine Wegstrecke von 10 cm verzögert eine Folge von Aktionspotentialen bereits um 1/10 Sekunde!
Theorem 1.9: Doppelte Negation neuronaler Signale
Werden erregende Signale in einem Umschaltkern zunächst auf hemmende Transmitter umgeschaltet und anschließend vermittels zweier hintereinander geschalteter Negationskerne doppelt negiert, so ist das Endsignal gegenüber dem Ursprungssignal
-einerseits zeitverschoben
-andererseits frequenzverändert.
Die Zeitverschiebung erfolgt einerseits
-durch die an jedem der drei beteiligten Neuronen auftretende Refraktärzeit, die nach einem abgefeuerten Signal vergehen muss, bevor ein neues Aktionspotential überhaupt entstehen kann
-durch die Endlichkeit der Ausbreitungsgeschwindigkeit der Aktionspotentiale, die sich entlang der beteiligten Axone und Dendriten zwischen Inputneuronen und Outputneuronen ausbreiten
-durch die Aufladungszeit, die nötig ist, bevor ein völlig entladenes Neuron durch eintreffende Signale seine Membranspannung so weitaufgeladen hat, dass es ein Aktionspotential abfeuern kann.
Die Frequenzverschiebung erfolgt, weil die Outputsignale mit der Einssignalfrequenz des zweiten Negationskerns feuern, diese entsteht aber durch Mittelwertbildung über dem einssignalbildenden Input und ist meist deutlich höher als die ursprüngliche Feuerfrequenz des Inputs.
Die Größe der Frequenzverschiebung hängt wesentlich von den Zeitkonstanten der beteiligten Neuronen ab sowie von der Frequenz der beiden Einssignalschwingungen der beiden Negationskerne. Grundsätzlich ist zu beachten, dass nur die Hüllkurven der beteiligten Signale zu beurteilen sind, da Negationskerne nur Hüllkurvensignale verarbeiten.
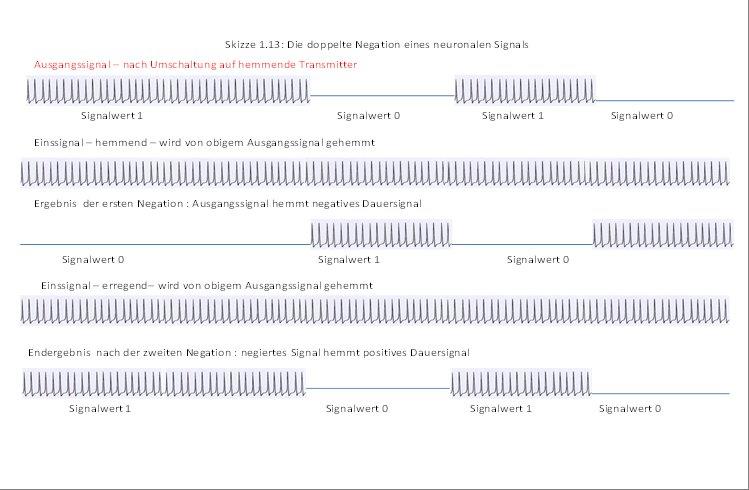
In der nachfolgenden Skizze 1.13 wird dargestellt, wie ein neuronales Signal, nachdem es auf einen hemmenden Transmitter umgeschaltet wurde, im ersten Schritt ein negatives (also hemmendes) Dauersignal (negatives Einssignal) hemmt. Das Ergebnis ist das negierte Ausgangssignal, welches ebenfalls hemmend ist.
Dieses hemmt nun im zweiten Schritt ein positives (erregendes) Einssignal (Dauersignal), wodurch das Endergebnis entsteht.
Vergleicht man die Hüllkurve des ursprünglichen Signals mit der Hüllkurve des durch doppelte Negation erzeugten Endsignals, so sieht man eine recht gute Übereinstimmung. Die Hüllkurven sind - bis auf eine eventuelle geringe Zeitversetzung - sehr ähnlich. Die ebenfalls mögliche Frequenzverschiebung konnte zeichnerisch nicht dargestellt werden.
Skizze 1.13: Die doppelte Negation eines neuronalen Signals (Hüllkurvensignals)
- ← Teil 1.2. Typisierung von neuronalen Kerngebieten
- Index
- Teil 1.4. Invertierung von neuronalen Signalen →
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan