Theorie der neuronalen Schaltung des Gehirns
und des analytischen Denkens
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
Teil 2.2. Die Rolle des Nucleus pedunculopontinus im Basalgangliensystem
Sowohl in Skizze 2.1 als auch in Skizze 2.2 zum Basalganglienmodell ist der Nucleus pedun-culopontinus Bestandteil des Systems. Dies trifft bereits beim zitierten Basalganglienschema von Young und Penney aus dem Jahre 2000 zu.
Eine nähere Analyse und vergleichende Betrachtungen führten den Autor auf die Ähnlichkeit zwischen Nucleus ruber und Nucleus pedunculopontinus:
- Beide Kerne benutzen erregende Einssignale. Der Nucleus ruber bildet diese selbst aus dem vom Cortex einströmenden Input der Mittelwertneuronen der vielen Cortexcluster, die zugehörigen Einssignalneuronen bilden seinen magnocellularen Anteil. Der Nucleus pedunculopontinus erhält sein externes Einssignal vom Nucleus subthalamicus.
- Beide Kerne empfangen hemmenden Input vom Globus pallidus interna des Striosomensystems (siehe Skizze 2.1).
- Beide Kerne erhalten hemmenden Input vom Globus pallidus externa des Matrixsystems (siehe Skizze 2.2).
- Beide Kerne erzeugen erregenden Output, jedoch ist der Output des Nucleus ruber glutamaterg, während der Output des Nucleus pedunculopontinus den erregenden Transmitter Acetylcholin verwendet.
Die erregenden Signale des Nucleus ruber erreichen über die Olive die Purkinjezellen als Kletterfasern. Der acetylcholinerge Output des Nucleus pedunculopontinus erreicht dagegen im Striosomensystem den Nucleus centromedianus des Thalamus (Skizze 2.1). Im Matrixsystem erreicht dieser Output auch den Thalamus, dieses Mal jedoch den Thalamus VA/VL.
Die Abkürzung VA steht für Nucleus ventralis anterior, VL steht für Nucleus ventralis lateralis. Beides sind Thalamuskerne.
Bekanntlich vertreten fast alle Neurologen die Lehrmeinung, der Thalamus wäre das Tor zum Bewusstsein, dessen Hauptsitz sich in der Cortexrinde befände. Insofern erstaunt es den Autor nicht, dass der im Basalgangliensystem mühsam erzeugte Output auch den Thalamus als erregendes Signal erreicht. Jedoch haben auch hier das Striosomensystem und das Matrixsystem unterschiedliche Zielpartner im Thalamus. Daher ist wiederum eine gewisse Aufgabenteilung zu vermuten.
An dieser Stelle sei nochmals an die Widmung im Kapitel 1 erinnert. Sie bezieht sich auf ein Gedankenmodell von Prof. Ulrich Ramacher. Dieser entwarf das Modell einer Auflösungspyramide. Wie sollte man sich diese vorstellen?
Ausgangspunkt für eine Erklärung könnte ein einfaches Bild sein. Wir denken uns also 1000 Bildpunkte in einer Reihe nebeneinander und 1000 solcher Bildpunktreihen untereinander. Daher hat das Bild eine Auflösung von 1.000.000 Pixeln, also eine Million Bildpunkte.
Wenn ein solches Bild eins zu eins in Neuronensignale umgesetzt würde, wären eine Million Rezeptoren mit zugehörigen Neuronen nötig. Diese Neuronen wollen wir als Startneuronen bezeichnen.
Nun kommen die Mittelwertneuronen ins Spiel. Stellen wir uns vor, ein Mittelwertneuron würde 25 der Startneuronen anzapfen und daraus einen gewichteten Mittelwert errechnen. Wir nennen diese Mittelwertneuronen die Mittelwertneuronen erster Stufe. Ihr rezeptives Feld beträgt 25 Pixel, also 5 Pixel Breite und 5 Pixel Höhe.
Damit wird die ursprüngliche Bildbreite von 1000 Pixel aber durch 5 dividiert, so dass wir in der Bildbreite nunmehr 200 Mittelwertneuronen erster Stufe wiederfinden.
Ebenso werden wir in der Bildhöhe statt 1000 Pixeln ebenso nur 200 Mittelwerte finden.
Verwenden wir nun anstelle der Startneuronen die Mittelwertneuronen, so entspricht dem Urbild mit 1.000.000 Pixeln ein Mittelwertbild erster Stufe mit 40.000 Pixeln. Dieses Bild hat also eine geringere Auflösung. Würden wir es wie das Urbild als Input zum Thalamus centromedianus senden, so hätten wir zwei Abbilder von der visuellen Welt: eines in Originalauflösung und ein „vergröbertes“, also auch „verkleinertes“ Abbild. Jedes Mittelwertneuron ermittelt also nicht nur die mittlere Aktivität im zugehörigen Cluster, sondern ist Outputlieferant für ein komprimiertes Abbild in der nächsten Stufe einer Abbildungspyramide. Nicht vergessen werden sollte, dass sich die Cortexcluster sehr wohl teilweise überlappen dürfen.
Skizze 2.3: Auflösungspyramide nach Ramacher als Bildzitat:
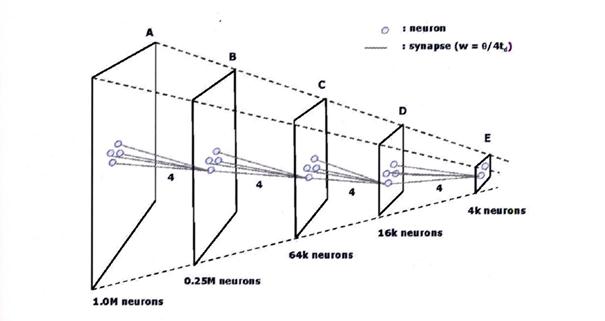
Quellenangabe: „Zur Konstruktion künstlicher Gehirne“, Ulrich Ramacher / Christoph von der Malsburg, 2009, Springer, Seite 323, Abbildung 16.5
Sind solche verkleinerten Abbilder nun Hirngespinste des Autors, reine Spekulationen oder Realitäten?
Ein derartiges Phänomen ist im Thalamus und in der Cortexrinde sehr wohl beobachtet worden. So schreiben Zilles/Rehkämper in der „Funktionellen Neuroanatomie“ vom Springer-Verlag auf Seite 230:
(Zitatbeginn)
„Der sekundäre somatosensorische Cortex S II enthält die Repräsentation der ipsa- und kontralateralen Körperseiten in geringer Ortsauflösung.“
„Das kleine, sekundäre Feld SII zeigt ebenfalls eine somatotope Repräsentation der Peripherie, wenn auch mit geringerer Ortsauflösung und auf der Grundlage einer weniger differenzierten architektonischen Gliederung. Die Verbindungen sind denen in S I grundsätzlich ähnlich, mit Ausnahme der Tatsache, dass hier eine Repräsentation beider Körperhälften gefunden wird, nicht nur der kontralateralen Seite wie in S I.“
(Zitatende)
Dieses Zitat aus der „Funktionellen Neuroanatomie“ von Zilles/Rehkämper möchte der Autor durch ein nachfolgendes Bildzitat aus dem überaus lesenswerten Werk „Das Gehirn“ von Richard F. Thompson vom Spektrum Akademischer Verlag ergänzen. Es ist der Abbildung 8.16 auf Seite 268 entnommen. Dem Verlag sei an dieser Stelle gedankt für die hervorragende Übersetzung des Originalwerkes aus dem Englischen. Ohne diese Übersetzung hätte der Autor sich niemals mit der Neurologie befasst, denn dieses Werk war das erste von ihm gelesene neurologische Fachbuch. Hätte es nur in Englisch vorgelegen, so wäre dieses Manuskript nicht entstanden.
Skizze 2.4: Bildzitat aus Thompson: Darstellung des Feldes S I und des zugehörigen, verkleinerten Feldes S II im somatosensorischen Cortex des Menschen
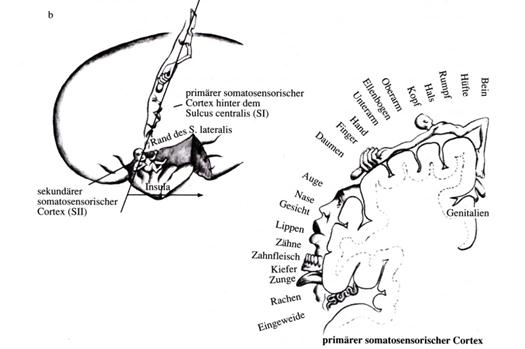
(Ende des Bildzitats)
Wichtig erscheint dem Autor der Hinweis, dass der Prozess der „verkleinerten Abbildung“ rekursiv ist, also ständig wiederholt wird, bis vom Urbild nur noch ein Mittelwert übrigbleibt. Dies liegt in der Annahme begründet, die Cortexrinde hätte überall eine ähnliche Grundstruktur. Wenn also der Nucleus centromedianus thalami die Mittelwertsignale der verkleinerten Abbildung zu einem neuen Cortexgebiet sendet, so werden auch dort wiederum Cortexcluster vorhanden sein. Diese beliefern inputmäßig neue Mittelwertneuronen der Schicht V, die ihrerseits über das Basalgangliensystem unter anderem wieder beim Nucleus centromedianus landen. So wiederholt sich alles, bis letztendlich nur noch ein Mittelwertneuron übrigbleibt.
Wir fassen unsere Erkenntnisse in einem neuen Theorem zusammen. Als Inhaber der Urheberrechte an dieser Erkenntnis benennt der Autor dieses Theorem nach dem Inspirationsgeber, Prof. Ulrich Ramacher.
Theorem 2.8: Theorem von Ramacher
Die Projektion des Nucleus pedunculopontinus zum Nucleus centromedianus ist Teil eines rekursiven Systems einer Auflösungspyramide, die vom Originalabbild rekursiv neue, verkleinerte Abbilder mit geringerer Auflösung produziert. Die Grundlage dafür bilden die Mittelwertneuronen von Schicht V der Cortexrinde, die über das Striosomensystem letztlich wieder den Input für ein vergröbertes Abbild zum Thalamus centromedianus senden, von wo aus es den Cortex erreicht.
Damit wäre die erregende Projektion des Striosomensystems (Skizze 2.3) vom Nucleus pedunculopontinus (7) zum Thalamus centromedianus (16) und anschließend zum Cortex geklärt.
Es verbleibt jedoch noch die erregende Projektion des Matrixsystems (Skizze 2.2) vom Nucleus pedunculopontinus (7) zum Thalamus VA/VL und von dort zum Cortex offen. Welche Signale sind es, die der NPP dem Cortex über den Thalamus zukommen lässt.
Hier erinnern wir uns an die Ähnlichkeit zwischen dem Nucleus ruber und dem Nucleus pedunculopontinus. Der Nucleus ruber sendet nach Theorem 2.5 den vierfach negierten Output der Signale der Signalneuronen des zugehörigen Cortexclusters (1) zur Olive, wo aus ihnen Kletterfasersignale werden.
Der Nucleus pedunculopontinus übernimmt, genau wie der Nucleus ruber, die letzte der vier Negationen. Denn bis zur Substantia nigra pars reticulata (6) der Skizze 2.2 sind die Ursignale des Cortex insgesamt dreifach negiert worden. Nun werden sie im Nucleus pedunculopontinus ein viertes Mal negiert und als erregendes Signal zum Thalamus VA/VL geleitet, von wo aus sie eine neue Cortexregion erreichen.
Damit empfängt nicht nur das Cerebellum das vierfach negierte Ursprungssignal der cortikalen Signalneuronen, sondern ebenfalls der Cortex (in einem neuen Projektionsgebiet).
Wir erinnern daran, dass das vierfach negierte Signal inzwischen stark zeitverzögert und frequenztransformiert ist. Also auch der Cortex erhält ein Abbild aller „Signale der Vergangenheit“, die etwa 125 Millisekunden zurückliegt.
Theorem 2.9: Zeitverzögerte Projektion der Signale der Signalneuronen eines Cortexclusters in ein neues Cortexgebiet
Die Projektion des Nucleus pedunculopontinus über den Thalamus VA/VL zu einem neuen Cortexcluster ist so organisiert, dass der Output eines jeden Signalneurons des Ursprungsclusters den neuen Cluster als Input erreicht, wobei das Signal im Matrixsystem vierfach negiert, zeitverzögert und frequenztransformiert wird. Die Zeitverzögerung hat in etwa die gleiche Größenordnung wie die Zeitverzögerung der gleichen Signale beim Weg zum Cerebellum über den Nucleus ruber und die Olive, jedoch sind letztere Signale die Kletterfasersignale des parvocellularen Systems.
In Teil 3 und Teil 4 dieser Monografie wird gezeigt, wie der Output des NPP genutzt wird, um zeitliche Veränderungen der Signallage, vornehmlich von Bewegungen von visuellen und auch motorischen Objekten (z. B. Gelenken) zu erkennen und zu digitalisieren, so dass sie im Cerebellum gespeichert und wieder abgerufen werden können. Dann wird auch erklärt, welche Aufgabe die millionenfachen Verbindungen der Cortexgebiete untereinander haben. Vorausgesetzt, die zugehörige Theorie erweist sich bei Prüfung als fehlerfrei.
Sowohl der Cortex als auch das Cerebellum erhalten also die durch das Matrixsystem zeitverzögerten Signale der Signalneuronen des Cortex. Da die ursprünglichen Cortexsignale zunehmend an systemtheoretischer Bedeutung gewinnen, wollen wir diesen Signalen einen weiteren, systemtheoretisch relevanten Namen geben.
Definition 2.2: Elementarsignale und Komplexsignale
Das Signal eines Signalneuronen eines Cortexclusters bezeichnen wir als Elementarsignal. Jedem Signalneuron Nk des Clusters entspricht also ein Elementarsignal, welches durch die Angabe der Feuerrate fk und seines Index k eindeutig beschrieben werden kann.
Ein Signalvektor S, dessen Komponenten die Feuerraten sämtlicher Elementarsignale des Clusters enthält gemäß
S = ( f1, f2, f3, ..., fn),
repräsentiert ein Komplexsignal K des Clusters.
Im Allgemeinen wird ein Komplexsignal als „Momentaufnahme“ betrachtet, da sich die Feuerraten seiner Komponenten zeitlich ändern können.
Nunmehr
können wir eine Analyse der Signale und Ausbreitungswege vornehmen.
- ← Teil 2.1. Das parvocellulare Kletterfasersystem des Gehirns
- Teil 2.3. Basalganglienerkrankungen und die tiefe Hirnstimulation →
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan