Vertebrate brain theory
ISBN
978-3-00-064888-5
Monograph of Dr. rer. nat. Andreas Heinrich Malczan
3.14 The origin of the primitive Pontocerebellum
In those Bilateria that were able to produce an original Spinocerebellum, a Pontocerebellum formed in parallel. This initially consisted of an extremely small number of neurons, but its formation took place according to a law.Let's recap: There were two types of output neurons in the early forebrain of this segmented bilateria. On the one hand, class 5 neurons projected downwards via the nucleus ruber to the motoneurons and via the nucleus olivaris to the primordial spinocerebellum. Those class 5 neurons that transmitted the vestibular signals projected to the vestibulocerebellum, the others to the spinocerebellum.
However, there were also neuron class 6 of the mean neurons of the first, cortical level. These projected directly into the thalamic mean centre (nucleus subthalamicus) and indirectly via the class 5 projection neurons to the mean centres of the lower floors.
Via the nucleus ruber, the cortical mean value signals also reached lower-lying mean value nuclei, so that the latter could supplement their segment-related mean values with these and thus also form body mean values. This descending projection was performed via the nucleus ruber.
Here the incoming signals in the nucleus ruber were treated equally. No matter whether the signals were from the vestibular receptors, the muscle tension receptors or average signals, all were treated equally (over time): They moved along the axons of nucleus ruber to the nucleus olivaris. The axons of the olivaris also moved to the developing cerebellum, which was initially a pure switch core (nucleus Purkinje) and switched the signals from glutamate to the transmitter GABA. In the cerebellum nucleus the inhibition of the permanently excited output neurons took place, which transmitted the excited output to the contralateral nucleus ruber. The permanent excitation originated from the reticular format. This corresponded to a signal inversion.
The original Cerebellum thus inverted exactly three types of signals:
- Vestibular signals
- Signals of muscle tension receptors (and other trunk receptors)
- Mean value signals
Similar signals formed their own partial structures in the cerebellum. From the part that inverted the vestibular signals, the vestibulocerebellum developed. Its cerebellar nucleus was the nucleus fastegii.
The part that initially inverted mainly the muscle tension receptors became the spinocerebellum. And since the motor signals came from two input nuclei, the nucleus cuneatus and the nucleus gracilis, the cerebellar nucleus of the spinocerebellum also divided into two sub-nuclei, the nucleus emboliformis and the nucleus globosus.
The third part, which inverted the average signals, became the original Pontocerebellum. His cerebellar nucleus became the nucleus dentatus.
It may seem presumptuous to speak of vestibulo-, spino- and pontocerebellum at such an early stage of evolution. These structures consisted of a collection of a few neurons. On the other hand, the Pontocerebellum as a whole certainly did not develop overnight, but must inevitably have undergone an evolution that started with a very small cluster of neurons. This monograph shows the further evolutionary path, as it can be reconstructed from current knowledge.
Theorem of the primitive Pontocerebellum
The primitive Pontocerebellum inverted the class 6 average signals of the uppermost cortical level. Its output was used for inverse excitation of the contralateral mean value systems, which it reached via the nucleus ruber.
Via the primeval Pontocerebellum the mean value systems exchanged their excitations.
The inverse excitation of the contralateral mean nuclei was topologically ordered by segments.
As in the early vestibulo- or spinocerebellum, there was no moss fibre projection and no granule cells in the primitive Pontocerebellum. Likewise there were no star cells, basket cells or golgi cells. The bark of the cerebellum consisted only of Purkinje cells, the surface of which we initially called the Purkinje nucleus. Their dendrite trees were certainly small and round in the beginning. The formation of inhibitory interneurons certainly began when lateral neighbor inhibition for contrast enhancement became a standard program in neuronal nuclei. However, such interneurons require an exciting signal influx, which was not yet present in the original cerebellum. For this purpose, an excitatory moss fibre projection had to be established first.
We assume that the mean signals already occupied a separate, coherent area in the nucleus olivaris, but due to the extremely low number of signals, this area only formed a very narrow strip that ran from the head side to the tail side and was present twice in the bilateral organism. Thus, the worm-like nucleus olivaris, if you imagine it arranged horizontally, got two narrow lateral stripes that represented the mean areas where the axons of the mean neurons arrived.
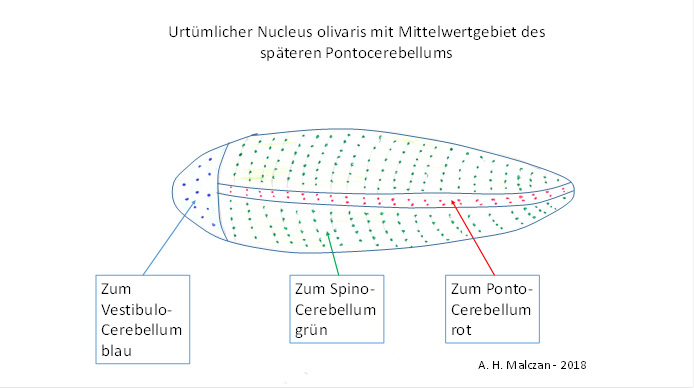
Figure 14 - Original nucleus olivaris
Since these neurons projected into the cerebellum, the same topological change took place there. The spinocerebellum, arranged horizontally, analogously formed an elongated, zeppelin-like body with two lateral strips. The axons arriving from the nucleus olivaris with the cortical mean signals ended in these strips, separately for each half of the body. These two strips formed the original Pontocerebellum. At the top of this structure were the neurons whose input came from the vestibular sense and were inverted in the associated vestibulocerebellum.
Many millions of years later, this elongated structure of the nucleus olivaris inflated, as did the associated worm-shaped cerebellum structure.
Significantly, this structure is actually called the worm of the cerebellum, in Latin Vermis cerebelli. If you think of the worm in a horizontal arrangement, its mean areas initially also formed two very narrow strips that ran from front to back on the left and right. The reason for this was the tendency to place neurons of the same type in a coherent fashion. In the front was the bilateral, relatively small and flattened vestibulocerebellum, also bilaterally present on the left and right.
And because the mean areas later expanded much more than the central parts belonging to the cerebellum worm, the bilaterally existing pontocerebellum developed laterally to the left and right of these. Since these are exactly the mean regions, we are entitled to call this early precursor structure in the original Spinocerebellum already here Pontocerebellum.
Parallel to the formation of the primordial Pontocerebellum, the signal classes 5 and 6 in the early basal ganglion system were treated equally in the early primordial brain of Bilateria, whose lines led to the vertebrates. Just like the signals of class 5, the signals of class 6 - via the nucleus subthalamicus - descended to the nucleus ruber. From there, the class 5 signals moved to the motor neurons, but the class 6 signals moved to the different mean nuclei.
Theorem of projections of the subthalamic nucleus
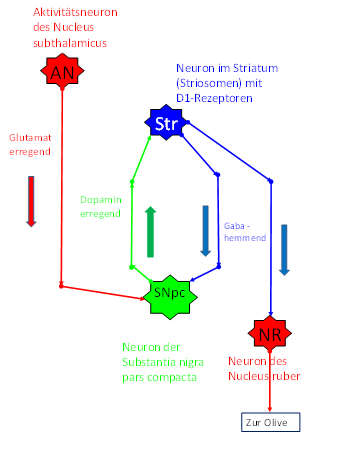
The output of the nucleus subthalamicus reaches the nucleus ruber in descending order, on the one hand, and the substantia nigra pars compacta and, via the cerebellum, the formatio reticularis on the other.
There was an exchange of signals between the mean systems of the different floors, with the nucleus subthalamicus being the highest ranking system. It also projected excitingly into the dopaminergic substantia nigra pars compacta.
Since each mean value system excitably projects back into the original regions of origin of the input, there is also an ascending projection from the substantia nigra pars compacta headwards. The mean value signals of the cortical clusters originating from the nucleus subthalamicus were switched to dopamine in the substantia nigra pars compacta and sent back, they ended in the striatum at GABAergic neurons, which are called striosomes. The striosome neurons are excited by this input because they use the dopamine receptor D2.
This led - due to the relatively large distance between the two structures - to a time delay of the striosome signals. The output of class 6 striosomal neurons reaches the nucleus ruber via the tractus tegmentalis centralis. There, there are already signal-related neurons that also receive the mean value signal of an assigned cortex cluster from the nucleus subthalamicus. The striatal axons of class 6 striosomes dock to these neurons and inhibit them with a time delay.
This resulted in a time-sensitive differential mapping of the cortical signal averages in the nucleus ruber. The striosome system was now able to detect temporal changes in signal class 5 as well as those in signal class 6.
Theorem of the time-sensitive difference mapping of the signal mean values in the nucleus ruber and the climbing fiber input of the pontocerebellum
In addition to signal class 5, signal class 6 of the mean value signals was also integrated into the striosome system, resulting in a time-sensitive differential mapping of the signal mean values in the nucleus ruber.
The equal treatment of the signals in the nucleus ruber led to the fact that these output signals of the mean-difference mapping reached the cerebellum via the nucleus olivaris. Since they were assigned to signal class 6, these signals reached the Purkinje cells of the forming pontocerebellumon the axons of the climbing fibres.
Thus, the primitive Pontocerebellum was able to detect temporal changes in the signal averages of the different segments of the body and to derive from them an inverse excitation of the contralateral segment parts. This made it possible to develop original forms of clock generators in the individual segments, which allowed asynchronous, opposing control of the motor functions and enabled meandering locomotion, as can still be observed in fish today.
We can now also divide the striosomes into two subspecies. One part processes the signals of signal class 5, the other the signals of class 6, which entitles us to speak of class 5 and 6 striosomes.
Theorem of classification of striosomes
The class 5 striosomes process class 5 signals and serve to form a time-sensitive differential mapping of the muscle tension receptors in the nucleus ruber, which sends them to the motor neurons as well as to the spinocerebellum via the nucleus olivaris. In the vestibulocerebellum it allows the detection of changes in body position in the earth's gravitational field, which are evaluated in the vestibulocerebellum. In the spinocerebellum, it allows the detection of signal changes of the different modalities and thus the detection of movement using different trunk and head senses.
The class 6 striosomes process the class 6 mean value signals and are used in the nucleus ruber to form a time-sensitive differential mapping of the mean value signals of the system. It allows the detection of temporal changes in the signal strength of the mean value systems. The nucleus ruber passes this output on to other mean value systems as well as via the nucleus olivaris to the Pontocerebellum.
In higher mammals, the axons from the striosomes to the nucleus ruber form a prominent nerve cord known as the tractus tegmentalis centralis. Initially, when the body of the animals had only a few muscles, it was extremely inconspicuous; the number of neurons involved was initially very small. Nevertheless, this neuron strand is to be called by this name already here.
However, with the inclusion of signal class 6 in the striosomes of the basal ganglia system and had to solve a related problem. The signal class 6 corresponded to the cortical mean values. Now there are enough cortical areas which are continuously supplied with input. Let us only think of the visual cortex. During the day, the visual cortex constantly receives signals from the visual receptors of the retina. Thus, the average signals formed from this are permanently strong and deliver a strong average signal to the substantia nigra pars compacta. This projects excitatory signals to the striatum.
However, a constant and strong permanent excitation is fatal for neurons. It was therefore advisable to dampen it. The solution was simple and obvious: the axons (of the mean signals) moving down from the striatum arrived at the nucleus ruber, where they caused the formation of the time-sensitive differential mapping. However, they continued on their way and also reached the substantia nigra pars compacta. There they had an inhibitory effect due to their transmitter GABA and prevented uninterrupted permanent excitation. Since the signals were strong average signals of class 6, their inhibitory effect was strong and stopped the continuous excitation in the substantia nigra pars compacta for several milliseconds until the GABAergic signals from the striatum were used up. Then the inhibition ceased and the substantia nigra was again excited by the cortical mean signals, which constantly reached this nucleus. Therefore, the substantia nigra could excite the striatum, or more precisely its striosomes. However, the striosomes projected back into this nucleus in an inhibitory manner, so that - due to the long distances - the inhibitory action potentials inhibited this nucleus again after a while.
Thus, the output of the substantia nigra was interrupted by regular inhibition pauses. Signal class 6 was transformed into a tonic (higher frequency) signal with regular pauses.
This is exactly what is known as the climbing fibre signal, which was detected in the Purkinje cells of the Pontocerebellum. It is exactly at these Purkinje cells when the assigned cortical mean neuron of class 6 is active, because in the corresponding catchment area of this mean neuron the cortex neurons are excited.
Theorem of inhibitory feedback from the striosomes of the striatum to the substantia nigra pars compacta
The projection of the cortical mean neurons of class 6 into the substantia nigra pars compacta required a limitation of excitation. Therefore, an inhibitory projection of the striosome neurons of the striatum into the substantia nigra pars compacta was established. This ensured that these neurons were not permanently and continuously excited, but that their tetanic oscillation was interrupted by short inhibition pauses during which the neurons could reactivate. The reason for the inhibition pauses was the transit time of the signals between cortex and substantia nigra pars compacta. These inhibition pauses were also impressed on the class 6 striosomal neurons. Their output signals inhibited the corresponding tetanic signals of class 6 in the nucleus ruber, so that the output signals of these neurons of the nucleus ruber were also tetanic oscillations, but interrupted by pauses. This output reaches the Pontocerebellum via the nucleus olivaris as climbing fiber signals.
The time delay explains the peculiarity of the climbing fibre signals in the Pontocerebellum, which consists in the fact that the higher frequency (tetanic) oscillation is interrupted by short pauses, which are important for the learning process.

Figure 15 - Climbing fiber
signal generated in the striosome system
Figure
16 - Dopaminergic and
GABAergic Projection in the Basal Ganglion System
The time delay of the dopaminergic signals on their way from the substantia nigra pars compacta to the striatum and the GABAergic signals from the striatum back to the substantia nigra pars compacta leads to periodic interruption of the mean value signals. Thus, the class 6 striosome system produces a characteristic oscillation form, which we (in this monograph) refer to as climbing fiber signal. In this way, each individual cortex cluster delivers its own climbing fiber signal to its cerebellum cluster in the pontocerebellum. Since the term cortex cluster has not been explained so far, for the sake of brevity it should be noted that a cortex cluster may represent the catchment area of a cortical mean neuron. This will be discussed in more detail later.
Already at this early stage of evolution, the prerequisites for the later learning ability of the Pontocerebellum were thus created. The high-frequency (tetanic) neural oscillation necessary for long-term potentiation (LTP) or long-term depression (LTD) is generated in the striosome system of the early basal ganglia. This was already happening at a time when the Pontocerebellum was just beginning to form.
Theorem of striosomal climbing fiber signals of the Pontocerebellum
The striosomal climbing fiber signals of the primordial pontocerebellum were tetanic oscillations interrupted by short pauses, which were indispensable for the later occurring long-term depression (LTD) and long-term potentiation (LTP) in the pontocerebellum. Whenever a neuronal excitation occurred in any cortex cluster, it was excited by the responsible mean neuron and sent to the nucleus subthalamicus as well as to the substantia nigra pars compacta, triggering a clocked permanent oscillation, which we refer to as climbing fiber signal. This was periodically interrupted by negative feedback to avoid overloading the neurons involved. Via the tractus tegmentalis centralis, the class 6 striosome signal reached the nucleus ruber and inhibited the incoming class 6 average signal from the nucleus subthalamicus in a point-to-point mapping, so that blanking pauses were forced upon it. This pulsed climbing fiber signal of a cortical cluster reached the Purkinje cells of the forming Pontocerebellum via the nucleus olivaris and finally via the climbing fibers, as well as the output neurons of the nucleus dentatus. Due to the existence of tetanic oscillations, a learning algorithm could develop in the pontocerebellum, which was based on the phenomenon of long-term depression (LTD) and later also on long-term potentiation (LTP).
The learning algorithm is described in the following chapters. It could only emerge when the second expansion phase of the Pontocerebellum began, in which cerebellar interneurons and a signal influx via a moss fiber system could develop.
Important for understanding the learning processes in the developing Pontocerebellum is the synchronicity of the oscillation processes in the striosomal system of the basal ganglia, which can be observed in the substantia nigra pars compacta as well as in the striosomes and in the nucleus subthalamicus. In [5] on page 287, Zilles and Rehkämper describe the pacemaker-like activity of the large, dopaminergic neuron in the substantia nigra pars compacta.
Theorem of synchronicity of the oscillation processes in the striosome system of the basal ganglia
In the substantia nigra pars compacta, the pauses between the many neurons involved are synchronized.
Theorem of striosomes as mean representatives of the cortex clusters
Since each class 6 neuron represents the signal average of a cortical cluster that is switched to dopamine via the substantia nigra pars compacta and reaches a class 6 striosome neuron in the striatum, each of these striosome neurons also represents the signal average of an associated cortical cluster. The mean value oscillations are interrupted by synchronized pauses.
It should be remembered that there was already a striosome system in the primordial triatum in which the signals of signal class 5 were processed in the same way as the signals of signal class 6. We can distinguish these striosome neurons by assigning them the signal class that they process. Then there are class 5 and class 6 striosome neurons, the latter belonging to the mean value system. Both types are excited by dopaminergic input.
Monograph of Dr. rer. nat. Andreas Heinrich Malczan