Theorie der neuronalen Schaltung des Gehirns
und des analytischen Denkens
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan
Teil 1.6. Die neuronalen Elementarschwingkreise im Striosomensystem
In der Skizze 1.9 wurde der Globus pallidus interna als negativer Negationskern für die Striosomensignale dargestellt.
In der Darstellung gibt es für jedes Aktivitätsneuron eine erregende Verbindung zu einem dopaminergen Neuron der Substantia nigra pars compacta. Dieses Neuron projiziert erregend auf ein GABAerges Striosomenneuron, dessen Axon sich verzweigt. Ein Axonzweig beliefert den Globus pallidus interna, während der zweite Zweig zurück auf das Neuron in der Substantia nigra pars compacta projiziert.
Die GABAerge Projektion des Striatums auf die Substantia nigra pars compacta ist bisher (im Juli des Jahres 2011) theoretisch ungeklärt.
Diese Monografie zeigt erstmals eine sinnvolle Deutung der Aufgabe dieser Projektion.
Diese wird umso bedeutsamer, als sich jetzt die Entstehung des Kletterfasersignals und seine Aufgabe wissenschaftlich deuten und belegen lässt.
Aus der Physik ist bekannt, dass ein Signal, das in einem System entsteht, zu seinem Ursprungsort zurückgeführt werden kann. In diesem Falle spricht man von Rückkopplung.
In elektrischen Schwingkreisen entstehen so Schwingungen, die ohne Rückkopplung nicht möglich wären.
Daher postuliert der Autor auch hier die Entstehung einer Schwingung. Diese Schwingung möge als Systemtakt des zugehörigen Aktivitätsneurons interpretiert werden. Sollte die Schwingungsentstehung nachweisbar sein, könnte ein Cortexcluster als Elementarcomputer des Gehirns mit eigenem Systemtakt interpretiert werden. Das Striosomensystem wäre eine vielfache Parallelschaltung solcher Elementarcomputer.
Theorem 1.13: Elementarschwingkreise im Striosomensystem
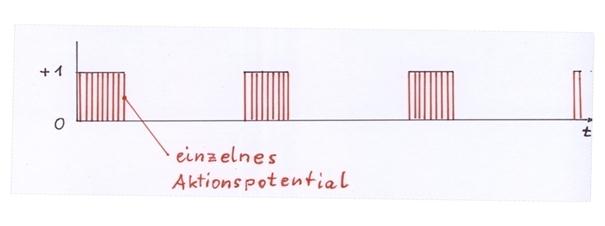
Jeweils ein Aktivitätsneuron eines Cortexclusters, das von ihm erregte dopaminerge Neuron in der Substantia nigra pars compacta und das von letzterem Neuron erregte GABAerge Neuron der Striosomen des Striatums bilden als Neuronentripel einen neuronalen Elementarschwingkreis. Dieser neuronale Elementarschwingkreis erhält als Input das Mittelwertsignal eines Cortexclusters. Im dem Falle, dass dieses Mittelwertsignal nicht das Nullsignal ist und längere Zeit andauert, transformiert der neuronale Elementarschwingkreis dieses zeitweilige Dauersignal in einen Systemtakt, der als Hüllkurvensignal ständig zwischen den Signalwerten 0 und 1 wechselt. Die Hüllkurvenfrequenz dieses neuronalen Systemtaktes beträgt ungefähr 5 Hz, so dass etwa 5 Schwingungen pro Sekunde auftreten, während die Feuerrate unterhalb der Hüllkurve ziemlich hoch ist.
Jeder Cortexcluster erzeugt im Falle der Aktivität des zugehörigen Aktivitätsneurons einen eigenen Systemtakt. Dennoch sind die Systemtakte der verschiedenen, benachbarten Cluster lose miteinander gekoppelt, was ein zeitweise synchrones Schwingen ermöglicht.
Es wird sich später zeigen, dass der neuronale Systemtakt eines Clusters zur Generierung eines Kletterfasersignals dient, welches der Abspeicherung der in diesem Cluster auftretenden Signalkombinationen dient.
Beweis zu Theorem 1.13:
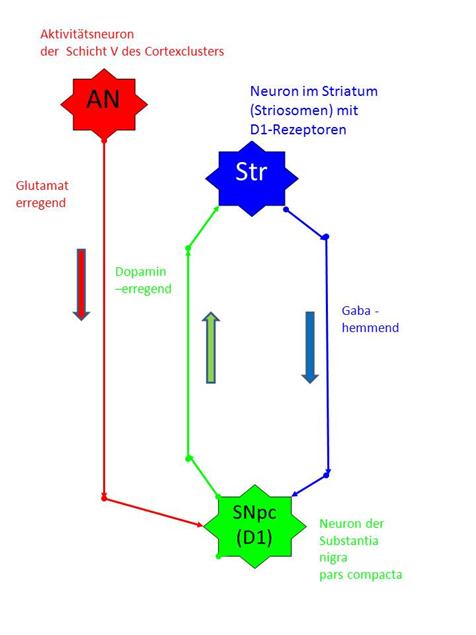
Wir zeichnen einen aus drei Neuronen bestehenden Elementarschwingkreis des Striosomensystems auf unter Beachtung der räumlichen Anordnung dieser Neuronenarten im Gehirn.
Oben befindet sich das Aktivitätsneuron AN der Schicht V des zugehörigen Cortexclusters, dargestellt in roter Farbe.
Unten befindet sich das Neuron SNpc der Substantia nigra pars compacta des D1-Systems, dargestellt in grüner Farbe.
Dazwischen befindet sich das zugehörige GABAerge Neuron STR des Striatums (blau).
International unbestritten ist die Tatsache, dass die Neuronen des Striosomensystems des Striatums der Basalganglien vermittels des Transmitters Gaba in Neuronen der Substantia nigra pars compacta projizieren. Dabei unterscheidet man in der Substantia nigra pars compacta zwei Klassen von Neuronen. Die erste Klasse, zu der unser Neuron SNpc gehören soll, projiziert unter Verwendung von Dopamin in die Striosomenneuronen, wobei der Rezeptortyp D1 durch Dopamin erregend wirkt (Subtyp D1).
Skizze 1.18: Elementarschwingkreise im Striosomensystem
Die zweite, hier zunächst vernachlässigte Klasse von Neuronen der Substantia nigra pars compacta projiziert mittels Dopamin in die Matrix des Striatums und wirkt dort über D2-Rezeptoren hemmend auf die Zielneuronen (Subtyp D2).
Wir zeichnen die drei zugehörigen Axone in obiges Schaltbild vom neuronalen Elementarschwingkreis ein:
Vom Neuron AN des Cortexclusters führt ein erregendes, glutamaterges Axon zum Neuron SNpc der Substantia nigra pars compacta vom Typ D1 (rot).
Vom Neuron SNpc der Substantia nigra pars compacta Subsystem D1 führt ein erregendes, dopaminerges Axon zum Neuron Str der Striosomen (grün).
Vom Neuron Str der Striosomen führt ein hemmendes, GABAerges Axon zum Neuron SNpc der Substantia nigra pars compacta Subsystem D1 (blau).
Unbestritten ist, dass die dopaminerge Projektion schwach myelinisierte Axone verwendet, wodurch sich die darauf befindlichen Aktionspotentiale langsamer ausbreiten.
Wir weisen die Entstehung des neuronalen Systemtaktes, der die Hüllkurve einer Folge von Aktionspotentialen sein möge, unter folgenden – nicht notwendigen – Voraussetzungen nach:
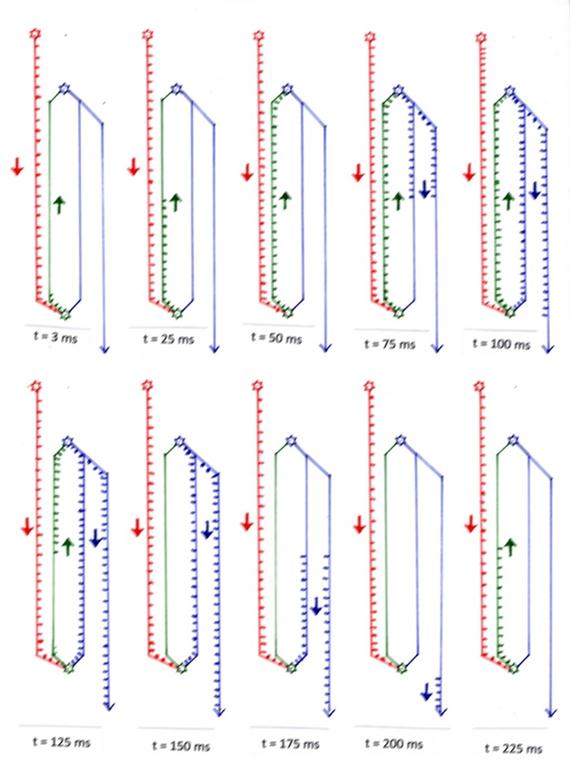
1. Der Abstand zwischen dem Neuron STR der Striosomen und des Neurons SNpc der Substantia nigra pars compacta betrage (beispielhaft) 50 mm.
2. Die Frequenz des Aktivitätsneurons AN betrage (beispielhaft) 200 Hz, so dass es 200 Aktionspotentiale pro Sekunde abfeuert.
3. Die Geschwindigkeit der Aktionspotentiale entlang der Axone betrage (beispielhaft) 1 Meter pro Sekunde, also 1 mm pro Millisekunde.
Zum Zeitpunkt t = 0 ms seien die zwei Neuronen STR und SNpc inaktiv. Am Neuron SNpc kommt das Dauersignal von Aktivitätsneuron AN bereits an. Jedes eintreffende Aktionspotential des AN-Neurons (rot) bewirke die Entstehung eines Aktionspotentials am SNpc-Neuron. Diese dort entstehenden, dopaminergen Aktionspotentiale breiten sich entlang des grün dargestellten Axons in Richtung des STR-Neurons aus. Nach t = 3 ms haben sie eine Strecke von 3 mm zurückgelegt, wie im ersten Bild der Zeitlupen-Darstellung ersichtlich ist. Zum Zeitpunkt t = 25 ms haben diese Aktionspotentiale, die ja noch immer laufend neu erzeugt werden, die Hälfte der Wegstrecke, also 25 mm, zurückgelegt (Bild 2).
Nach 50 Millisekunden (Bild 3) kommen die dopaminergen Aktionspotentiale endlich erstmalig am Neuron STR der Striosomen an. Dort erzeugen sie wegen ihrer erregenden Wirkung wiederum jeweils ein Aktionspotential, welches sich nun in umgekehrte Richtung zum SNpc-Neuron bewegt. Nach 75 Millisekunden hat dieses gabaerge Aktionspotential bereits die Hälfte des Rückweges hinter sich gebracht. Zum Zeitpunkt t = 100 ms erreichen die GABAergen, also hemmenden Aktionspotentiale erstmalig das SNpc-Neuron und bewirken dort eine starke Hemmung, die mindestens für die kurze Zeit von 5 Millisekunden anhalten möge (Refraktärzeit).
Dadurch bleiben aber die bisherigen dopaminergen Aktionspotentiale aus, da das zugehörige SNpc-Neuron nunmehr dauerhaft gehemmt bleibt, solange bei ihm hemmende Aktionspotentiale eintreffen. Zum Zeitpunkt t = 125 ms schweigt das SNpc-Neuron bereits 25 Millisekunden, während noch Aktionspotentiale für die Hemmungsdauer von 75 Millisekunden auf den Schwingkreisaxonen (blau und grün) zur Verfügung stehen.
Nach 150 Millisekunden sind alle Aktionspotentiale des dopaminergen Axons aufgebraucht, aber das gabaerge Axon enthalt noch hemmende Aktionspotentiale für genau 50 Millisekunden bereit. Diese sind zum Zeitpunkt t = 125 ms erst zur Hälfte aufgebraucht.
Genau zum Zeitpunkt t = 200 ms kommt das letzte hemmende, gabaerg wirkende Aktionspotential am SNpc-Neuron an und hemmt es letztmalig. Danach sind alle bisherigen Aktionspotentiale des blaugrünen Weges aufgebraucht, und eine evtl. noch vorhandene Erregung des cortikalen Aktivitätsneurons AN kann wieder ungehindert zur Erregung des SNpc-Neurons führen. Der Vorgang beginnt von neuem.
Zusammenfassend lässt sich feststellen:
1. Für die Zeitdauer von 100 ms ist das Neuron der Substantia nigra pars compacta erregt und sendet Aktionspotentiale aus.
2. Für die Zeitdauer von 100 ms ist das Neuron der Substantia nigra pars compacta gehemmt und sendet keine Aktionspotentiale aus.
3. Für die Zeitdauer von 100 ms ist das Neuron der Striosomen des Striatums erregt und sendet Aktionspotentiale aus.
4. Für die Zeitdauer von 100 ms ist das Neuron der Striosomen des Striatums gehemmt und sendet Aktionspotentiale aus.
5. Zwischen SNpc-Neuron und STR-Neuron besteht hinsichtlich Erregung und Hemmung eine Phasenverschiebung (Zeitverschiebung) von 100 ms.
6. In dem dargestellten System aus drei Neuronen existiert eine neuronale Schwingung, die für die angegebenen Zahlenbeispiele eine Frequenz von 5 Hz besitzt, also 5 Schwingungen pro Sekunde besitzt. Diese Schwingung ist die Hüllkurve der beteiligten Aktionspotentiale. Die Aktionspotentiale unter der Hüllkurve haben die Frequenz von 200 Hz, wie zu Beginn des Beweises (beispielhaft) vorgegeben wurde.
Zeitlupendarstellung der Schwingungen im Striosomenschwingkreis
Skizze 1.19: Schwingung im Striosomensystem – der neuronale Systemtakt eines Clusters
Theorem 1.14: Lose Synchronisation der Schwingungen im Striosomensystem
Die vielen Einzelschwingkreise in System Globus pallidus interna, Substantia nigra pars compacta (Subklasse D1) und in den Striosomen sind durch lose Kopplung in der Substantia nigra pars compacta teilweise synchronisiert.
Beweis zu Theorem 14: Wörtliches Zitat aus „Funktionelle Neuroanatomie“ Zilles/Rehkämper, 3. Auflage, Springer Verlag Berlin Heidelberg 1998, Seite 287, erster und zweiter Absatz, mit freundlicher Genehmigung des Autors
(Zitatbeginn)
„In der Substantia nigra ist ein sehr zelldichter Bereich, Pars compacta, und ein Bereich mit geringer Zellpackungsdichte, Pars reticularis, erkennbar. Während die Pars reticularis der Substantia nigra GABAerge Neurone besitzt, die auf die motorischen Thalamuskerne projizieren, enthält die Pars compacta überwiegend große dopaminerge Neurone, die eine langsame (2 – 5 Hz) schrittmacherartige Aktivität aufweisen und deren Axone das Striatum erreichen.
Neben dem Gehalt an Melanin, das der Substantia nigra die namensbestimmende schwarze Farbe verleiht, weisen dopaminerge Neurone mehrere Besonderheiten auf. Sie zeigen synchrone Aktivität, die auf dendrodendritische Synapsen zwischen den Neuronen und dendritische Freisetzung von Dopamin zurückgeführt wird. Die Freisetzung von Dopamin an den Axonterminalen wird stärker durch präsynaptische Rezeptoren als durch Schrittmacheraktivität reguliert.“
(Zitatende)
An dieser Stelle sei den Herausgebern, Professor Karl Zilles sowie Professor Gerd Rehkämper für die überaus präzise Darstellung der Cytoarchitektonik speziell der Basalganglien gedankt, ebenso Herrn Professor Rolf Kötter. Besonders die systematischen Beschreibungen zur Cytoarchitektonik der Basalganglien durch Professor Rolf Kötter, die noch heute im Internet verfügbar sind, waren bei der Erarbeitung dieser Theorie die inspirationsliefernden Elemente.
- ← Teil 1.5. Die mengentheoretische Betrachtung von neuronalen Kernen
- Teil 1.7. Die Erzeugung des (magnocellularen und primären) Kletterfasersignals ... →
ISBN
978-3-00-037458-6
ISBN 978-3-00-042153-2
Monografie von Dr. rer. nat. Andreas Heinrich Malczan